





(C) 2011 S.V. Pavlova. This is an open access article distributed under the terms of the Creative Commons Attribution License 3.0 (CC-BY), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
For reference, use of the paginated PDF or printed version of this article is recommended.
We report a new finding of the 54-chromosome sibling species of the common vole in East Siberia - the first description from Olkhon Island (Lake Baikal). The karyotype of a male specimen revealed by routine staining and C-banding demonstrates the unambiguous presence of Microtus rossiaemeridionalis Ognev, 1924 (recently often regarded as as junior synonym of Microtus levis Miller, 1908). Comparison with conspecific specimens from the European part of the species range (from the left bank of the river Volga) shows that the vole of the island population has a smaller X-chromosome due to a reduced quantity of C-positive heterochromatin. This is just the third example of this type of X-chromosome variant with previous cases on an Arctic island (Svalbard) and the West Siberian lowland (Novosibirsk) and the only one on a lake island. Although Microtus rossiaemeridionalis is largely monomorphic in its karyotype, our data show that one specific type of X-chromosome variant is remarkably widespread, though rare.
chromosome sibling species, common voles, Microtus arvalis group , Microtus rossiaemeridionalis, Lake Baikal, X-chromosome
The investigation of intraspecific variability of chromosomes is one of the traditional approaches to study evolutionary processes. Comparative karyological investigations of new and especially extreme localities of widely distributed species are of particular interest (
Since the description of two sibling species in the common vole differing in diploid numbers (2n=46 in Microtus arvalis Pallas, 1779 and 2n=54 in Microtus rossiaemeridionalis Ognev, 1924) (
Here we report one of the easternmost findings of the 54-chromosome karyotype for Microtus rossiaemeridionalis, from an isolated population on Olkhon Island in Lake Baikal.
Materials and methodsSmall mammal trapping was conducted in July 2008 in Irkutsk province, East Siberia, on the western shore of Lake Baikal and on Olkhon Island that was separated by a narrow channel (2-5 km in width) from the mainland. A sole adult male of the common vole was live-trapped on Olkhon Island about 800 m from the village of Khalgai (53°42'14"N; 107°31'32"E) at the edge of larch – pine forest, bounded by steppe habitat.
For the cytogenetic comparison, fresh chromosome preparations were prepared in a similar way for 3 specimens, 2 males and 1 female, collected in March 2011 in the European part of the range of the common vole (village of Dyakovka on the left bank of the river Volga, Saratov province: 50°42'54"N; 46°45'52"E). These animals were caught using live-traps on the bushy slope of the right bank of the river Yeruslan, about 400 m from the village and about 2.5 km from the Dyakovsky Forest (Fig. 1). In addition to our data, a recent Siberian collection site (Novosibirsk, West Siberia) is indicated in the distribution map of the Microtus rossiaemeridionalis taken from the official web-site of A.N. Severtsov Institute of Ecology and Evolution (http://www.sevin.ru/vertebrates/).
Materials for cytogenetic and further molecular analyses were fixed in the field following a standard protocol (
Standard mitotic and meiotic chromosome preparations were obtained in the field from the bone marrow and from testes following
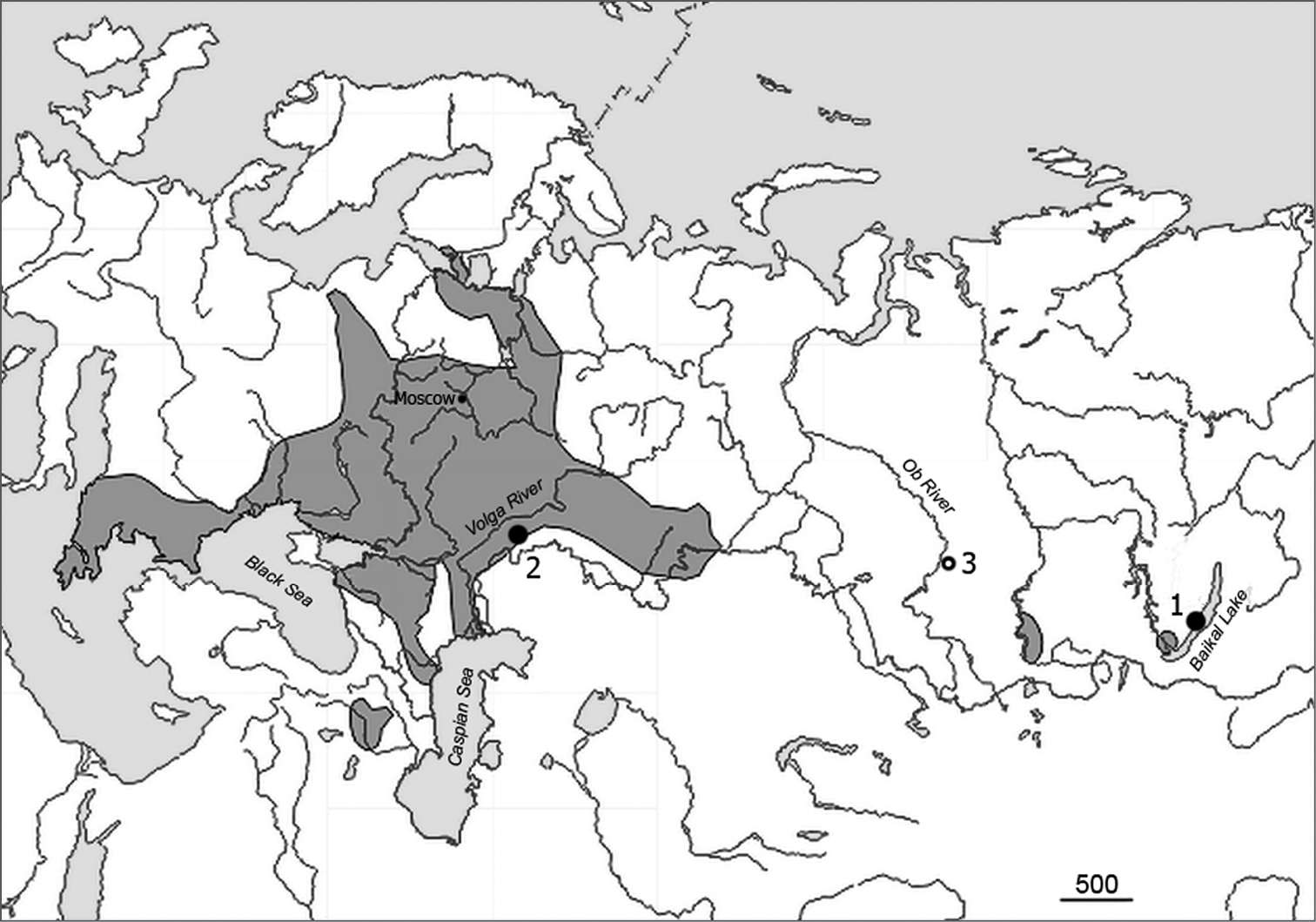
Map showing collection sites for the 54-chromosome (Sibling) vole under study. Black circles – our data (1 Olkhon isl., Lake Baikal 2 Dyakovka, left bank of Volga River.) Open circle – most recent Siberian finding 3 Novosibirsk vic., right bank of Ob River. Colored are the territories where verified findings (karyotype, allozymes) were obtained from.
The four voles examined had the same chromosome number 2n=54 and identical autosomal karyotype supplemented by the typical sex chromosomes complement – XX in females, XY in males. All chromosomes but the smallest pair of metacentric autosomes were acrocentric (NF=56). Routine staining was ineffectual in identifying more than two pairs of large acrocentrics and the small metacentric pair from the morphologically homogeneous group of medium to small acrocentrics. After C-banding, the centromeres of all autosomes were positively C-stained. The two largest elements with additional C-blocks of heterochromatin were classified as the pair of sex chromosomes, the Y being totally heterochromatic and the X carrying a large telomeric block (Fig. 2). The X chromosome was always the largest element in the complement and the Y the next largest, but close in size to the largest autosome.
However, the length of the X chromosome varied between voles from the two geographically distant regions. In the male from Olkhon Island both sex chromosomes looked alike in routinely stained karyograms and did not exceed considerably in size the largest autosome (Fig. 2a). They differed each from the other only by C-banding, and in this case the distal heterochromatic block marking the X occupied less than a half of its total length (Fig. 2b). Similar proportions in length of the sex chromosomes were seen in meiotic plates of this individual showing X and Y stick configuration. Autosomes formed bivalents during meiosis whereas sex chromosomes remain asynaptic (Fig. 3).
In stark contrast, in voles from the European sample the X was larger than either the Y or the larger autosomes, and this can only be due to a larger amount of telomeric heterochromatin occupying the distal half of the X chromosomes in males (Fig. 2c) as well as in a female studied.
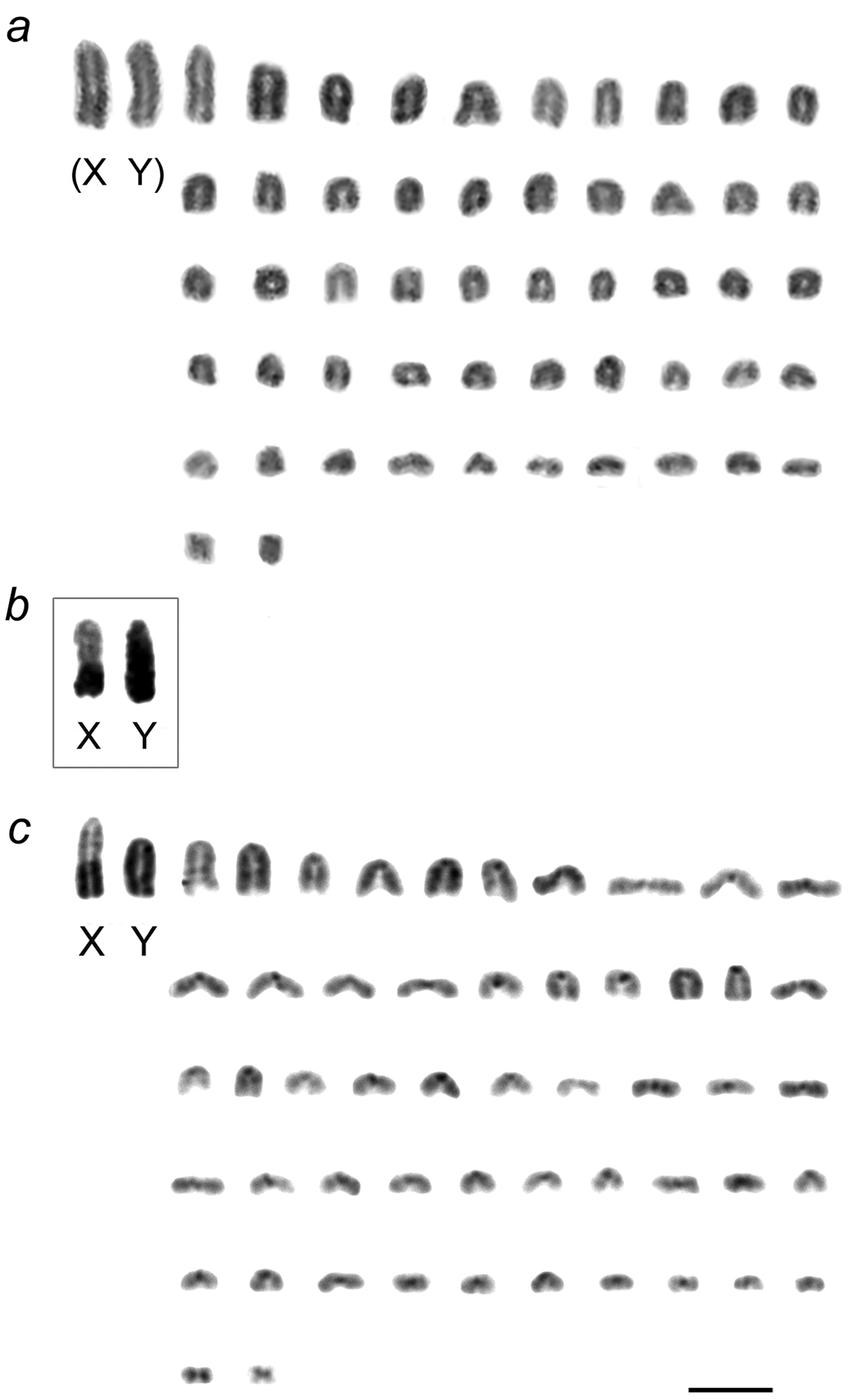
Mitotic chromosomes of East Siberian and East European Microtus rossiaemeridionalis: aconventionally stained chromosomes of the male from Olkhon Island arranged by size, with provisional identification of the XY sex chromosome pair, based on chromosome length b C-banded sex chromosomes of the same individual c C-banded chromosomes of a male from the east bank of the river Volga. Bar=10 µm.
The chromosomal characteristics of the specimens studied are consistent with the karyotypic features of the 54-chromosome sibling species of the common vole, a member of the Microtus arvalis group (
It is interesting that although the ranges of the two sibling species, Microtus arvalis and Microtus rossiaemeridionalis, significantly overlap in Eastern Europe and in their Asian parts; these species have been found, in general, to be separated in Siberia and Kazakhstan (
Further chromosomal studies added only a few occurrence sites for Microtus rossiaemeridionalis in Kazakhstan (
Meiotic preparations in a male from Olkhon Island (Fig. 3) revealed that the sex chromosomes remain asynaptyc which is typical for Microtus rossiaemeridionalis and the related species (
Cytogenetically, our findings indicate that in the Microtus rossiaemeridionalis karyotype, which otherwise is being considered rather stable, there is an X chromosome predisposition to intraspecific variation. Our data indicate that the variation affects the heterochromatic part of the X chromosome and have shown the value of karyotypic investigations on new and especially extreme localities in uncovering new karyotypic variability. Even in a rather invariant species like Microtus rossiaemeridionalis, such studies are worthwhile.
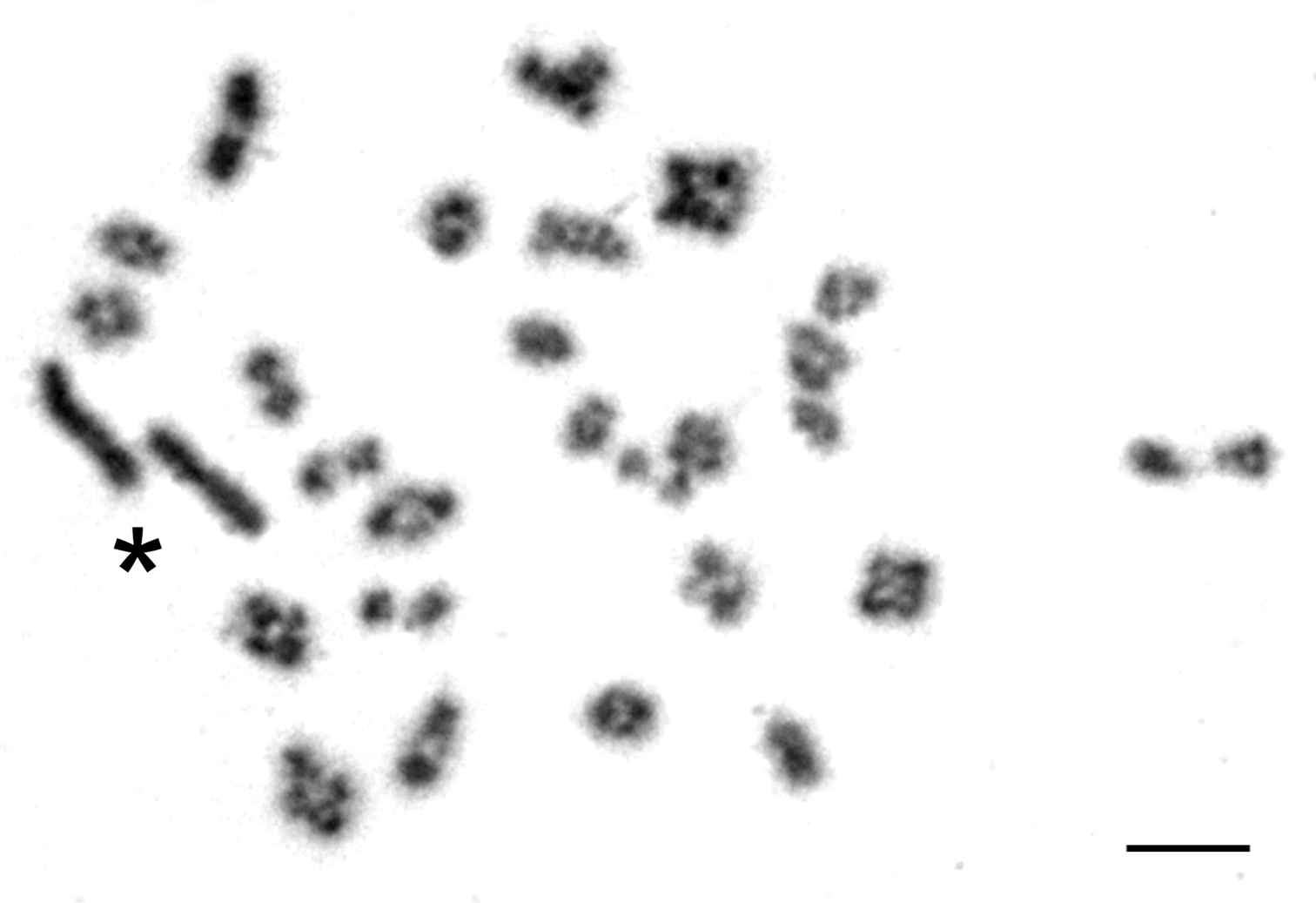
Meiotic spread with 26 autosomal bivalents and the characteristic asynaptic configuration of the sex chromosomes (star) from a male common vole of Olkhon Island (2n=54). Bar=10 µm.
We greatly appreciate the assistance of L. Savinetskaya, A. Kriukov and other colleagues, who participated in collecting data in the field. We thank Dr. Nina Sh. Bulatova who stimulated this study. Dr. J.B. Searle read the earlier version of the manuscript and helped in improving English. Financially, this study was supported by the Federal Target Program “Scientific and scientific-educational manpower of innovative Russia” (contract 02.740.11.0282), the Program for Fundamental Research of Presidium of RAS “Biological diversity”, RFBR (11-05-01066) and the President Grant for Russian Distinguished Young Scientists MK-2500.2011.4.