





(C) 2012 Maressa Ferreira-Neto. This is an open access article distributed under the terms of the Creative Commons Attribution License 3.0 (CC-BY), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
For reference, use of the paginated PDF or printed version of this article is recommended.
Ninety individuals of the characid fish Astyanax fasciatus (Cuvier, 1819) were collected at Água da Madalena stream (Botucatu, São Paulo, Brazil) and analyzed for diploid chromosome number 2n and karyotype composition as well as for the chromosomal location of the 5S and 18S ribosomal DNA (rDNA). Whereas no chromosome differences were associated with sex, three different karyomorphs with diploid chromosome numbers 2n=46, 2n=48 and 2n=50 were found. No intermediate 2n numbers were discovered. The 2n=50 karyomorph showed some differences in 18S rDNA location compared to the two other karyomorphs. Finally, all specimens with the 2n=46 karyomorph showed the presence of a partly heterochromatic macro supernumerary chromosome, which was absent in all individuals with the two other karyomorphs. All these results suggest that indviduals of the three different karyomorphs are not likely to hybridize in the examined populations. Our findings strongly suggest the presence of three separate species (sensu biological species concept) easily diagnosed on the basis of differences in the diploid chromosome numbers and other chromosomal markers.
fish cytogenetics, chromosome banding, rDNA, sympatric differentiation, B chromosome
The genus Astyanax (Baird et Girard, 1854) is one of the most diversified among Neotropical characid fishes. Indeed, this genus shows an extensive morphological diversification and a highly complex taxonomy. Moreover, a number of species actually appear to be “complexes of species” with low morphological differentiation but high variation at other levels, e.g. chromosome number and morphology (
Astyanax fasciatus is another species that seems to form a “species complex” since the available information points towards the existence of several karyomorphs with the diploid chromosome numbers 2n=45, 2n=46, 2n=47, 2n=48 and 2n=50 (Table 1). The karyotype most frequently cited in the literature is 2n=48 (Table 1), but the known geographical range is actually larger for the karyomorphs 2n=50 and 2n=46 (Figure 1).
Diploid numbers (2n) recorded in populations of Astyanax fasciatus complex.
| Species as originally reported | River/basin | 2n | B chrom. | Reference |
|---|---|---|---|---|
| Astyanax fasciatus | Mogi Guaçu river/Paraná | 45 | 14 | |
| Astyanax fasciatus | Mogi Guaçu river/Paraná | 46 | 2, 4, 9, 10, 11, 18 | |
| Astyanax fasciatus | Mogi Guaçu river/Paraná | 47 | 14, 11 | |
| Astyanax fasciatus | Mogi Guaçu river/Paraná | 48 | 11, 18 | |
| Astyanax fasciatus | Piracicaba river/Paraná | 48 | 7 | |
| Astyanax fasciatus | Piracicaba river/Paraná | 46 | 23 | |
| Astyanax fasciatus | Tietê river/Paraná | 46 | 5 | |
| Astyanax fasciatus | Riacho Águas da Madalena/Paraná | 50 | 23 | |
| Astyanax fasciatus | Águas da Madalena stream/Paraná | 48 | 23 | |
| Astyanax fasciatus | Águas da Madalena stream/Paraná | 46 | + | 23 |
| Astyanax fasciatus | Paranapanema river/Paraná | 46 | 6 | |
| Astyanax fasciatus | Paranapanema river/Paraná | 50 | 13 | |
| Astyanax fasciatus | Paraíba river/Paraná | 48 | 3, 5, 9 | |
| Astyanax fasciatus | Paiol Grande river/Paraná | 48 | 9, 17 | |
| Astyanax fasciatus | Barra funda river/Paraná | 46 | 9 | |
| Astyanax fasciatus | Passa Cinco river/Paraná | 46 | 9, 11 | |
| Astyanax fasciatus | Sapucaí river/Paraná | 48 | 12 | |
| Astyanax fasciatus | Araguari river/Paraná | 46 | 16 | |
| Astyanax cf. fasciatus | Juquiá river/Paraná | 48 | 2 | |
| Astyanax fasciatus | Meia Ponte river/Araguaia | 46 | 1 | |
| Astyanax prope fasciatus | Córrego Fundo stream/Araguaia | 50 | 8 | |
| Astyanax fasciatus | Araras river/Paraná | 48 | 22 | |
| Astyanax fasciatus | Patos river/Paraná | 48 | 22 | |
| Astyanax fasciatus | Três Bueiros river/Paraná | 48 | 22 | |
| Astyanax fasciatus | Almas river/Paraná | 48 | 22 | |
| Astyanax prope fasciatus | Tibagi river/Paraná | 48 | 19 | |
| Astyanax prope fasciatus | Tibagi river/Paraná | 49 | 19 | |
| Astyanax prope fasciatus | Tibagi river/Paraná | 50 | 19 | |
| Astyanax fasciatus | São Francisco river/São Francisco | 48 | 7, 9, 17, 22 | |
| Astyanax fasciatus | São Francisco river/São Francisco | 46 | + | 7 |
| Astyanax fasciatus | Três Marias/São Franscisco | 46 | + | 15 |
| Astyanax fasciatus | Contas river/Leste | 48 | 21 | |
| Astyanax fasciatus | Mineiro do Costa stream/Leste | 48 | 21 | |
| Astyanax fasciatus | Preto do Costa river/Leste | 48 | 21 |
References: 1. Jim and Toledo (1975); 2.
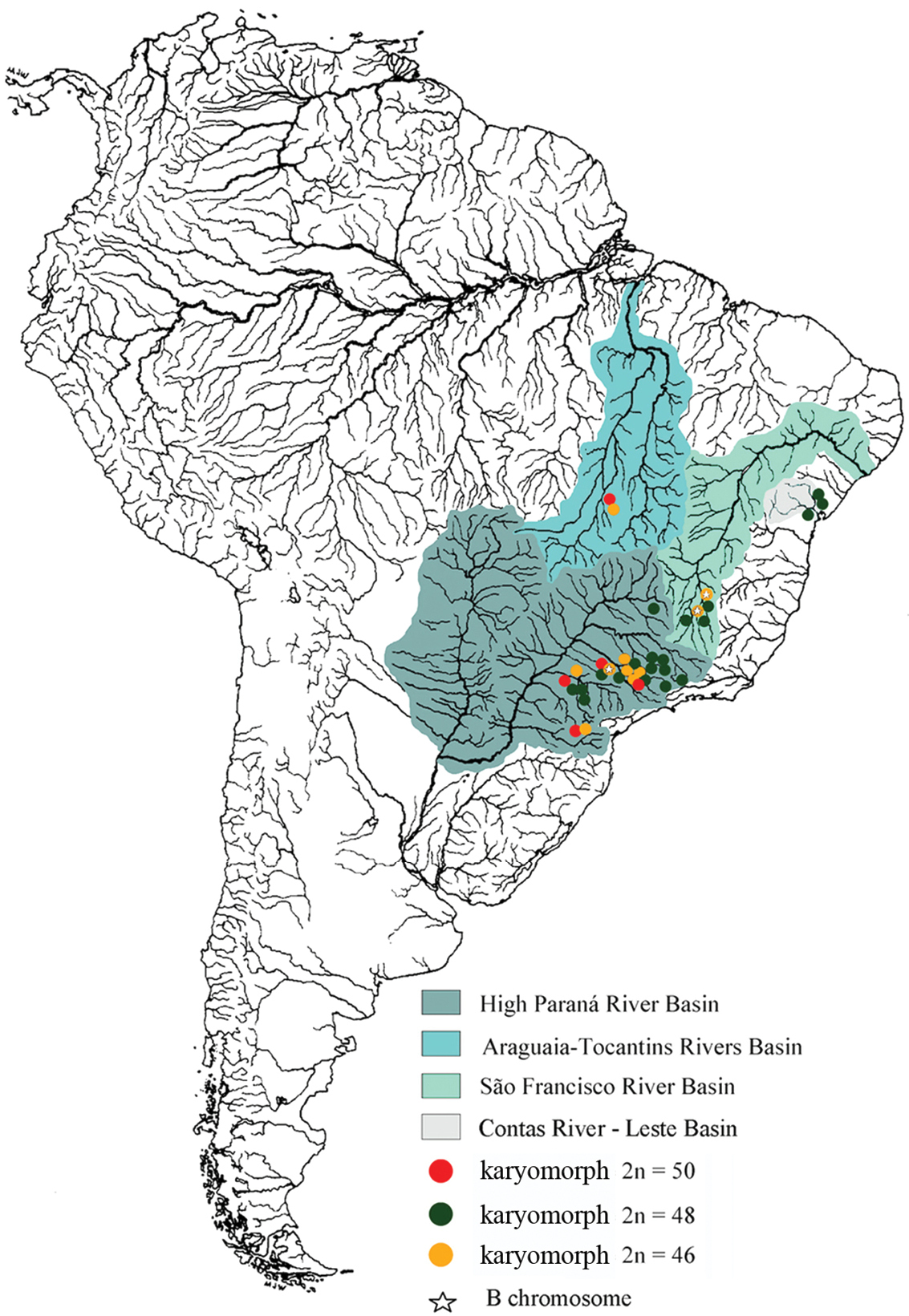
Map of South America highlighting the hydrographic basin of the Paraná, Araguaia-Tocantins, Contas and São Francisco Rivers. The natural distribution of Astyanax fasciatus and the presence of B chromosomes are represented according to the legends.
B chromosomes are supernumerary elements previously reported in the karyotypes of several Astyanax species (
In this study we analyzed the 2n=46, 2n=48 and 2n=50 Astyanax fasciatus karyomorphs by examining a high number of individuals living in sympatry. The absence of intermediate chromosome numbers, the restriction of B chromosomes to the 2n=46 karyomorph and the absence of heteromorphism for the 18S rDNA sites (which differ among some karyomorphs) strongly suggest that the abovementioned karyomorphs are not likely to hybridize and are thus three separate species (sensu biological species concept).
Material and methodsNinety Astyanax fasciatus specimens (56 females and 34 males) were collected at a 3.000 m2 area of the Água da Madalena stream, Botucatu, SP, Brazil (22°59'23"S, 48°25'31"W). The specimens were anaesthetized in benzocaine (1%) and, after collecting tissue samples for chromosome analysis, fixed in 10% formol and preserved in 70% alcohol. After identification, the specimens were deposited in the Museum of Biology and Laboratory of Fish Genetics, UNESP, Botucatu, São Paulo, Brazil. Chromosome preparations were obtained from anterior kidney cells and used for the conventional air drying technique (
Metaphase chromosomes were counterstained with DAPI and analyzed under optical light microscope (Olympus BX61). Images were captured using the Image-Pro Plus 6.0 software (Media Cybernetics). To build karyograms, chromosome morphology was determined according to the arm ratio limits established by
The cytogenetic analysis of the 90 specimens of Astyanax fasciatus revealed the presence of three different karyomorphs in the sample, showing 2n=46, 2n=48 and 2n=50 chromosomes (Table 2 and Fig. 2a, c and e, respectively), and with fundamental numbers (NF= number of chromosome arms) equal to 84, 86 and 90, respectively. The anatomical sex of the fish, determined by visual examination of the gonads, was not associated with the karyotypic differences, so that males and females were found with all the three cytotypes described here. Interestingly, all the specimens with the 2n=46 karyomorph showed the presence of one mitotically stable macro B chromosome (Fig. 2a, b), whereas the specimens of the 2n=48 and 2n=50 karyomorphs lacked it. No individual with intermediate odd chromosome numbers was found.
Cytogenetic studies in Astyanax fasciatus karyotype composition and location of the 5S and 18S rDNA in the chromosomes of the individuals analyzed.
|
Cytotypes 2n / NF |
Number of specimens |
Chromosome | rDNA 18S/5S |
Number of B Chromosomes |
NOR | |
|---|---|---|---|---|---|---|
| Female Male | Formulae | location (pairs) | females | males | (pairs) | |
| 2n = 46, NF=84 | 12 10 | 5m+8sm+6st+4a | 8, 15, 16, 20, 21, B/3, 20 | 1 | 1 | 9, 10 |
| 2n = 48, NF=86 | 10 9 | 5m+8sm+6st+5a | 8, 12, 19, 20, 21/3, 20 | 0 | 0 | 9, 10 |
| 2n = 50, NF=84 | 41 32 | 5m+8sm+4st+8a | 5, 8, 12, 18, 22/3, 21 | 0 | 0 | 9, 10 |

Karyotypes of Astyanax fasciatus (2n=46, 48 and 50 chromosomes) deduced after conventional Giemsa staining (a, c, e) and double FISH using 5S (green) and 18S rDNA (red) probes. The chromosomes bearing the Ag-NORs are boxed in b, d and f. Bar = 10 μm.
Physical mapping of the rDNA showed remarkable conservation of the 5S rDNA sites, which were present in two chromosome pairs, i.e. Nos. 3 and 20 (21 in the 2n=50 karyomorph) (Fig. 2b, d, f).
The silver staining technique revealed the presence, in all cells, of Ag-NORs on two sm chromosome pairs of the three karyomorphs. One Ag-NOR was located at the distal region of the p arm and the other at the q arm (Figs 2b, d, f, details). FISH analysis, however, showed the presence of 18S rDNA at the distal regions of five chromosome pairs, but no FISH signal was observed on the short arm of any chromosome (Fig. 2b, d, f). This points towards the presence of a small number of 18S rRNA genes in the short arm of the sm chromosome.
Double FISH showed the presence of a chromosome pair, No. 20 in the 2n=46 and 2n=48 karyomorph, carrying both 5S (proximal) and 18S (distal) rDNA, but no chromosome carried both kinds of rDNA in the 2n=50 karyomorph (Fig. 1f). All six specimens analyzed by FISH were homomorphic for the chromosome No. 20 which carries both rDNA types. Remarkably, FISH mapping showed the presence of an interstitial cluster of 18S rDNA in the long arm of the B chromosome found in the 2n=46 karyomorph. Nevertheless, this rDNA cluster was never detected by silver staining.
DiscussionSympatry and syntopy for several cytotypes have been reported in Astyanax species, such as Astyanax scabripinnis (Souza and Moreira-Filho 1995) and Astyanax fasciatus (
Our present analysis of a large sample of individuals caught in the same stream shows syntopic occurrence of three Astyanax fasciatus karyomorphs with discrete chromosome numbers 2n=46, 2n=48 and 2n=50. The absence of intermediate chromosome numbers, the presence of B chromosomes in only one of these karyomorphs (2n=46), and the absence of apparent heteromorphism for the chromosome 20, suggests that the three karyomorphs do not hybridize in this stream. In such case, the data point out towards the possibility that these three karyomorphs actually correspond to three cryptic species, thus supporting the hypothesis that Astyanax fasciatus is in fact a species assemblage, i.e. several species were included under the nominal name of Astyanax fasciatus (
The B chromosome found in Astyanax fasciatus is large and metacentric. Interestingly, both characteristics seem common to all the B chromosomes described Astyanax species including Astyanax scabripinnis (
Since silver staining of metaphasic chromosomes reveals only those NORs that were active in the previous interphase (
The higher number of rDNA clusters (10) than silver stained NORs (4) suggests the inactivity of some rDNAs in most cells. It would therefore be interesting to ascertain whether this phenomenon is facultative or constitutive. Silver nitra te may also bind to other proteins present in the nuclei, implying that some chromosome structures visualized by silver nitrate may not correspond to ribosomal genes (
The presence of one B chromosome in all 22 individuals from the 2n=46 karyomorph is intriguing, since it departs from the usual interindividual variation which characterizes B chromosomes (
This study was supported by the Fundação de Amparo à Pesquisa do Estado de São Paulo – FAPESP (Proc. 2008/57067-1), Conselho Nacional de Desenvolvimento Científico e Tecnológico – CNPq, and Coordenação de Aperfeiçoamento de Pessoal de Nível Superior – CAPES.