





(C) 2012 Valentina G. Kuznetsova. This is an open access article distributed under the terms of the Creative Commons Attribution License 3.0 (CC-BY), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
For reference, use of the paginated PDF or printed version of this article is recommended.
Using the fluorescence in situ hybridization (FISH), the presence of (TTAGG)n telomeric sequence was detected in the chromosomes of Lethocerus patruelis (Stål, 1854) belonging to the family Belostomatidae (Heteroptera: Nepomorpha). This sequence was exclusively present at the ends of chromosomes in this species. This is the first evidence of the insect-type TTAGG telomeric repeats in Heteroptera.
Chromosomes, FISH, (TTAGG)n telomeric repeat , true bugs, Nepomorpha, Belostomatidae, Lethocerus patruelis
Telomeres are specific nucleoprotein structures at the ends of chromosomes and are responsible for their stability. Information on the telomere structure and function is presently available for many animals, plants and fungi (
In this paper we report the molecular structure of telomeres at the physical ends of chromosomesin Lethocerus patruelis (Stål, 1854) (Nepomorpha: Belostomatidae).
Material and methodsSpread chromosome preparations were made from testes of Lethocerus patruelis and stained using a Shiff-Giemsa method as described in
At first metaphases in Lethocerus patruelis males, 11 autosomal bivalents, each with one (sometimes two) terminal or subterminal chiasmata, a bivalent of m-chromosomes (micro-chromosomes) and a XY- pseudo-bivalent could be seen (Fig. 1a). Figures 1b-d show the results of fluorescence in situ hybridization with pentanucleotide (TTAGG)n and 18S rDNA probes to several meiotic spreads. At metaphase nuclei, TTAGG fluorescent signals (red) are clearly seen at all chromosomal ends, whereas rDNA clusters (green) are clearly evident on the X and Y chromosomes (Fig. 1b, c). Prominent telomere clustering at the periphery of spermatid nuclei (Fig. 1d) creates one large while sometimes a small number of TTAGG signals (red).
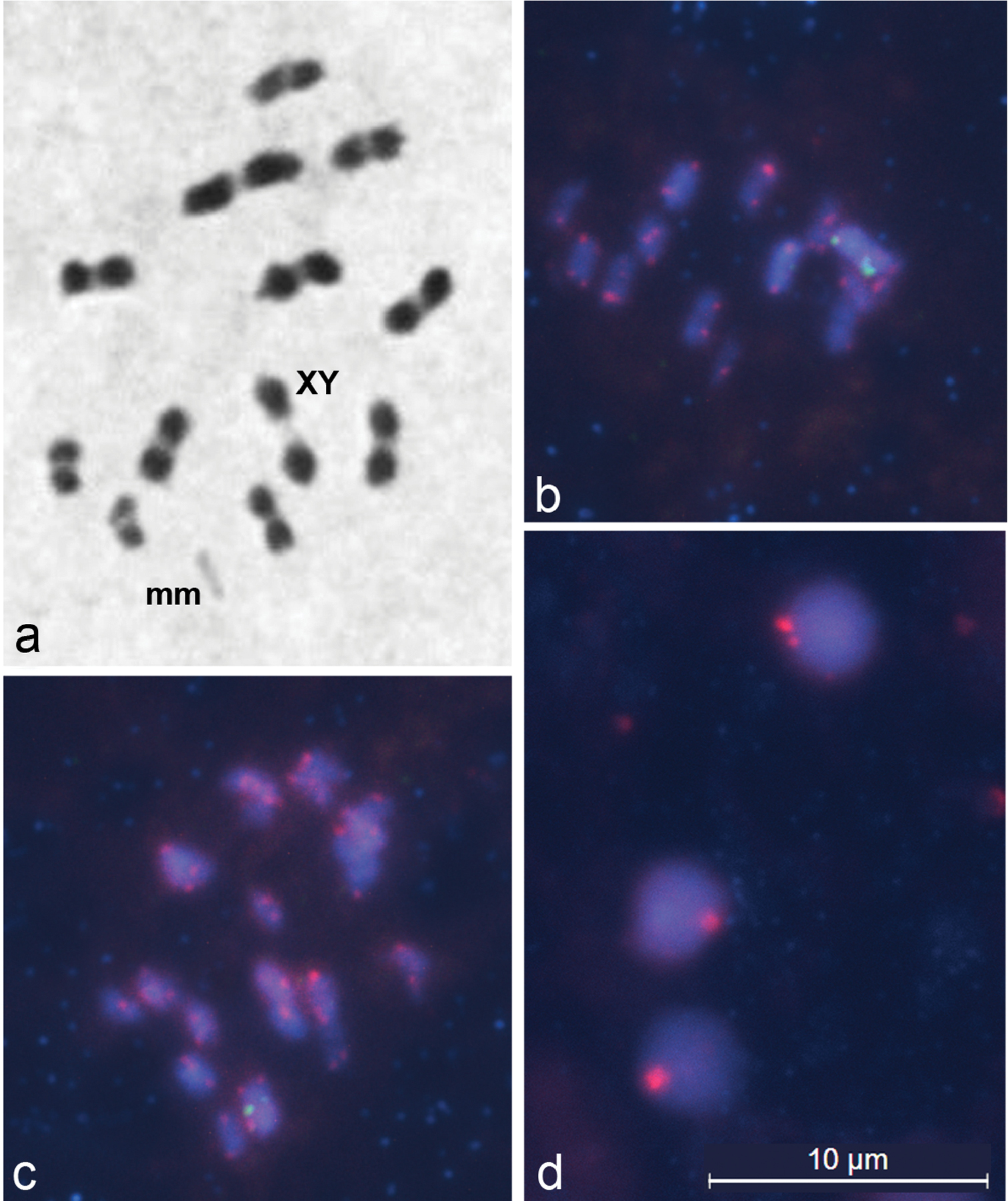
Meiotic chromosomes of Lethocerus patruelis subjected to standard staining (a) and FISH (b–d). a metaphase I showing n = 11AA + mm + XY; b–d representative FISH images of metaphase I chromosomes (b, c) and spermatids (d) hybridized with probes against 18S rDNA and telomeres, showing ribosomal clusters (green) on X and Y chromosomes (b, c), and TTAGG repeats (red) located at the ends of chromosomes (b, c) and clustered at the periphery of spermatid nuclei (d).
The standard karyotype of Lethocerus patruelis males is 2n = 22A + 2m + XY as it was recently shown by
DNA of the telomeres consists of short nucleotide motifs (combinations) repeated thousands and millions of times. Comparative analysis of these motifs in various groups of organisms has shown that they are evolutionarily stable, and, having once appeared during the evolution, mark taxa and phylogenetic lineages of high rank (
Quite recently,
We emphasize, however, that the problem of telomere composition in different insect orders is still not adequately explored and in most cases, the available data concern one or more species only (see Fig. 6 in
In Heteroptera, the absence of the (TTAGG)n telomeric motif was firstly shown for Halyomorpha halys (Stål, 1855) (Pentatomidae) studied using Southern hybridization (Okazaki et al. 1993: as Halyomorpha mista (Uhler, 1860)) and Pyrrhocoris apterus (Linnaeus, 1758) (Pyrrhocoridae) subjected to both Southern hybridization and FISH (
Originally proposed by
Our results of FISH with a (TTAGG)n probe strongly demonstrated that (TTAGG)n sequence was located at the telomeres of all chromosomes in Lethocerus patruelis. The finding of the insect-type (TTAGG)n telomeric motif in Lethocerus patruelis is thus clearly indicative of the heterogeneity of Heteroptera in telomere organization. The family Belostomatidae, to which this species belongs, is classified within the infraorder Nepomorpha (or true water bugs). The data on telomeres imply that true water bugs preserved the plesiomorphic telomere structure, whereas Cimicomorpha and Pentatomomorpha have the apomorphic state of this character, which can be considered a synapomorphy of these infraorders. This conclusion is consistent with the generally accepted opinion that Cimicomorpha and Pentatomomorpha represent a monophyletic lineage, and Nepomorpha has a basal position within Heteroptera (
The study was supported (for VK and BA) by the RFBR (grant 11-04-00734), programs of the Presidium of the RAS “Gene Pools and Genetic Diversity” and “Origin of the Biosphere and Evolution of Geo-biological Systems” and the Ministry of Education and Science of the Russian Federation, and (for SG) by DO-02-259 NSF Sofia, BAS.