





(C) 2013 Adauto Lima Cardoso. This is an open access article distributed under the terms of the Creative Commons Attribution License 3.0 (CC-BY), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
For reference, use of the paginated PDF or printed version of this article is recommended.
The family Loricariidae encompasses approximately 800 species distributed in six subfamilies. The subfamily Hypostominae consists of five tribes; of them, the tribe Ancistrini is relatively diverse, but it is not well known from the cytogenetic point of view. Genus Scobinancistrus Isbrücker et Nijssen, 1989, which is part of the tribe Ancistrini, has two species that occur in sympatry in the Xingu River, Brazil. In this work, we performed the first karyotypic characterizations of these two species and sought to identify the processes involved in their karyotypic evolution. Chromosomal preparations were subjected to Giemsa staining, silver nitrate impregnation, C-banding, CMA3 staining, DAPI staining, and FISH (fluorescence in situ hybridization) with 18S rDNA and telomeric probes. We found that Scobinancistrus aureatus Burgess, 1994 and Scobinancistrus pariolispos Isbrücker et Nijssen, 1989 shared the diploid number, 2n=52, but differed in their karyotypic formulae (KFs), distribution of constitutive heterochromatin (CH), and the localizations of their nucleolus organizer regions (NORs), which were found on the interstitial and distal regions of the long arm of chromosome pair 3 in Scobinancistrus aureatus and Scobinancistrus pariolispos respectively. We suggest that these interspecific variations may have arisen via paracentric inversion or transposition of the NOR. The karyotypic differences found between these two Scobinancistrus species can be used to identify them taxonomically, and may have functioned as a mechanism of post-zygotic reproductive isolation during the speciation process.
Karyotypic divergence, chromosome rearrangements, sympatry
The fishes of the family Loricariidae are an important component of the ichthyofauna in the Neotropical region, where they are widely distributed and occupy a great variety of freshwater environments (
Cytogenetic information is incipient, given the great diversity of Ancistrini species, and has been efficient in distinguishing species of this tribe (
The genus Scobinancistrus Isbrücker et Nijssen, 1989 (Hypostominae, Ancistrini) comprises two species: Scobinancistrus pariolispos Isbrücker et Nijssen, 1989, which occurs in the Tapajós, Xingu and Tocantins Rivers, Brazil, and Scobinancistrus aureatus Burgess, 1994, which is endemic to the Xingu River (
In the present work, we obtained Scobinancistrus pariolispos and Scobinancistrus aureatus from the Xingu River and studied their karyotypes, in an effort to identify the processes involved in their karyotypic evolution and contribute new cytogenetic information for members of the tribe Ancistrini.
Samples of Scobinancistrus aureatus (seven females) and Scobinancistrus pariolispos (five males and two females) from the Xingu River, Brazil, were analyzed (Fig. 1). Metaphase chromosomes were obtained according to the method described by Bertollo et al. (1978) and analyzed by conventional staining (Giemsa), C-banding (Sumner 1972), Ag-NOR labeling (
The specimens of Scobinancistrus aureatus and Scobinancistrus pariolispos obtained from the Xingu River both had diploid numbers 2n=52 chromosomes, but they differed in their karyotypic formulas (KFs), which were 22m-20sm-10st and 24m-18sm-10st, respectively (Fig. 2A, B). Both males and females were analyzed for Scobinancistrus pariolispos, but no sex chromosomes were identified.
C-banding failed to identify constitutive heterochromatin (CH) in the centromeric region of any chromosome in the studied species. In Scobinancistrus aureatus, large heterochromatic blocks were seen, as follows: in the proximal regions of the long arms of chromosome pairs 5, 6 and 18; in the proximal regions of the short arms of pair 13; throughout the short arms of pair 22; in the distal regions of the long arms of pairs 3 and 12; and in the distal regions of the short arms of pairs 12 and 18 (Fig. 2C). In Scobinancistrus pariolispos, conspicuous heterochromatic blocks were also identified throughout the short arms of pair 2, in the distal regions of the long arms of pair 3, and throughout the long arms of pair 5 (Fig. 2D).
A NOR was identified on a single chromosome pair per species. In Scobinancistrus aureatus, the NOR was located in the interstitial region of the long arms of pair 3, flanked by CH (Fig. 2E). In Scobinancistrus pariolispos, the NOR was situated in the distal region of the long arms of pair 3, adjacent to a block of CH (Fig. 2F). The numbers and localizations of these regions were confirmed by FISH with 18S ribosomal DNA probes (Fig. 3E, F).
The DAPI fluorochrome labeled the heterochromatic regions (Fig. 3A, B), and CMA3 stained the NORs (Fig. 3C, D). In FISH using telomeric sequence probes (TTAGGG), the probes hybridized to the ends of all chromosomes, but no interstitial telomeric labeling was observed (Fig. 3G, H).
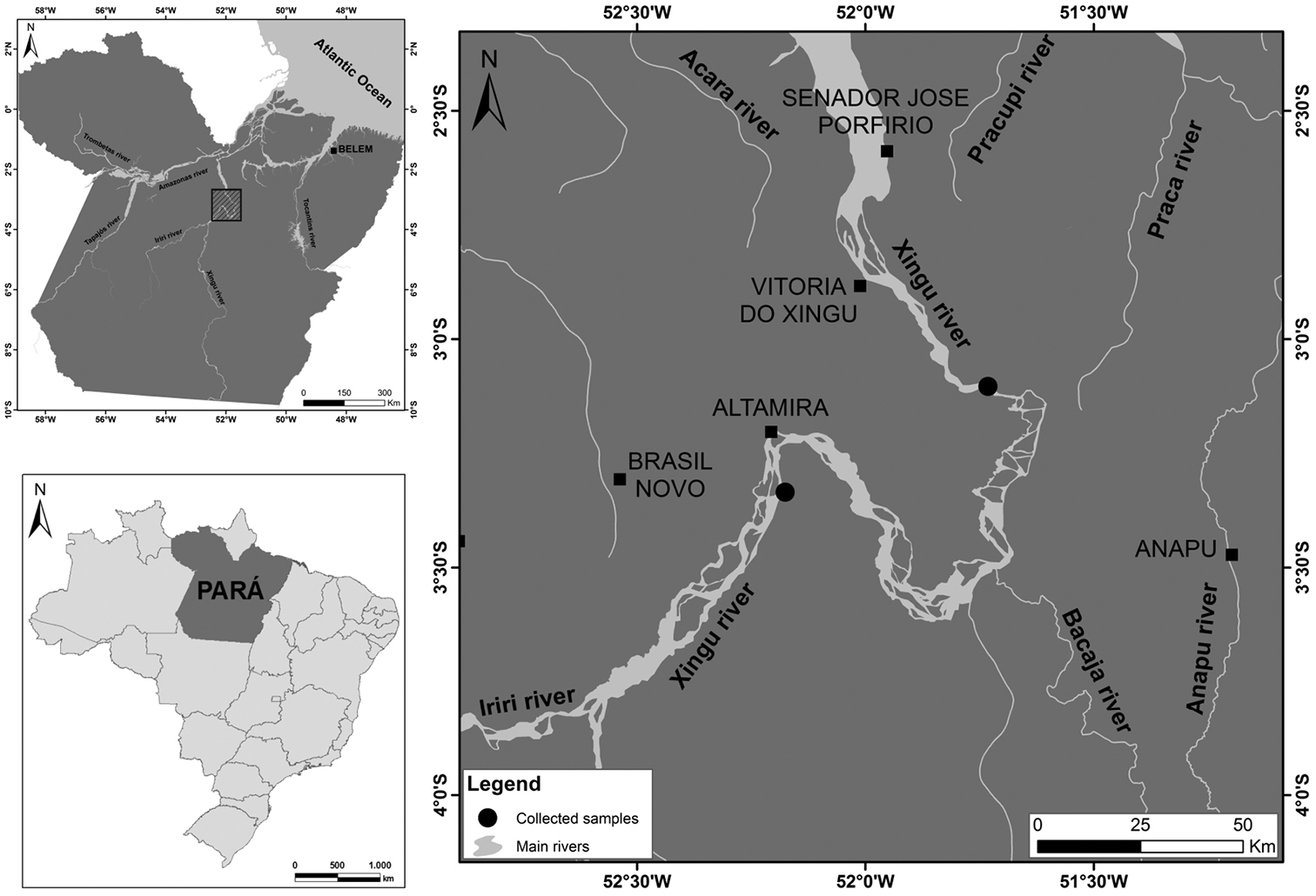
Collection localities of the analyzed Scobinancistrus samples.
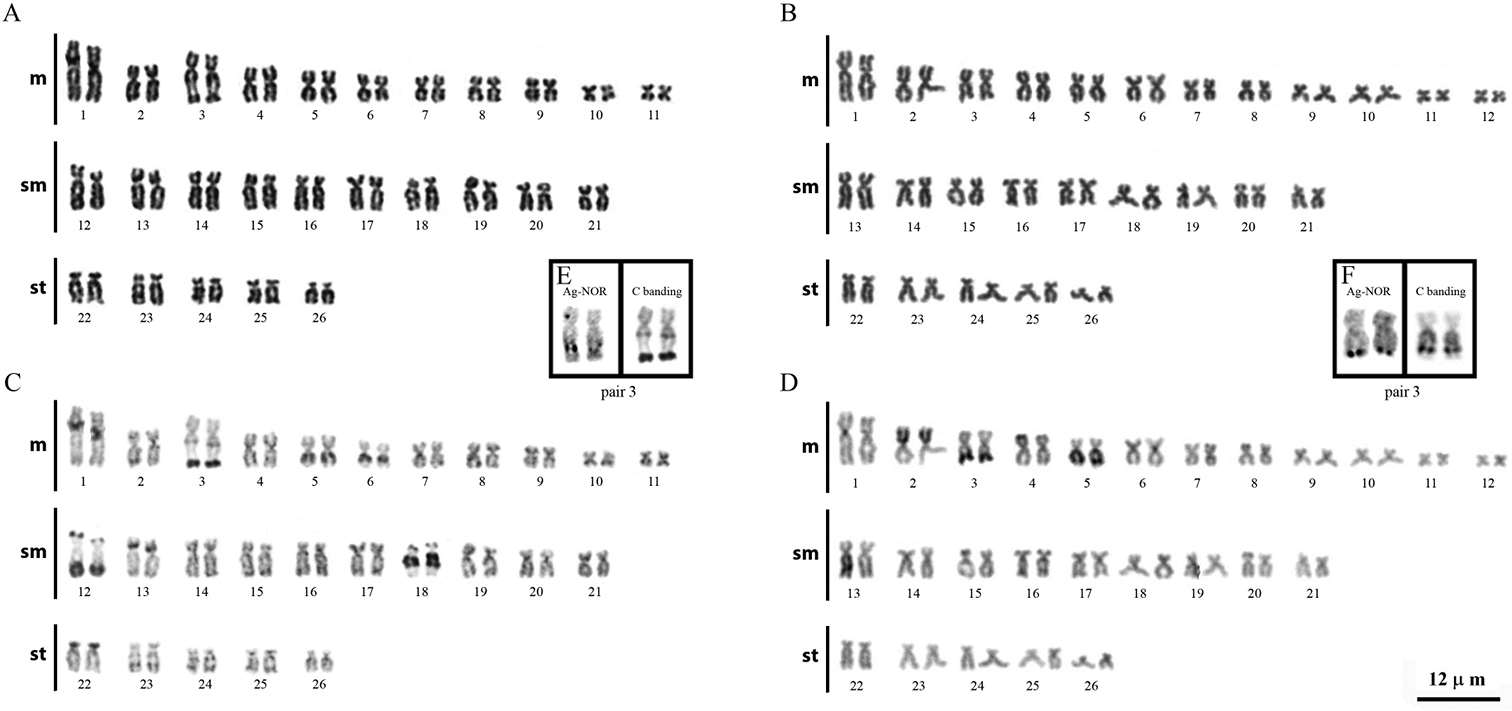
Karyotypic analyses of Scobinancistrusaureatus (A, B and E) and Scobinancistrus pariolispos (B, D and F): conventional staining (A and B), C-banding (C and D) and the NOR-bearing chromosome pair (E and F). The scale bar refers to all images.
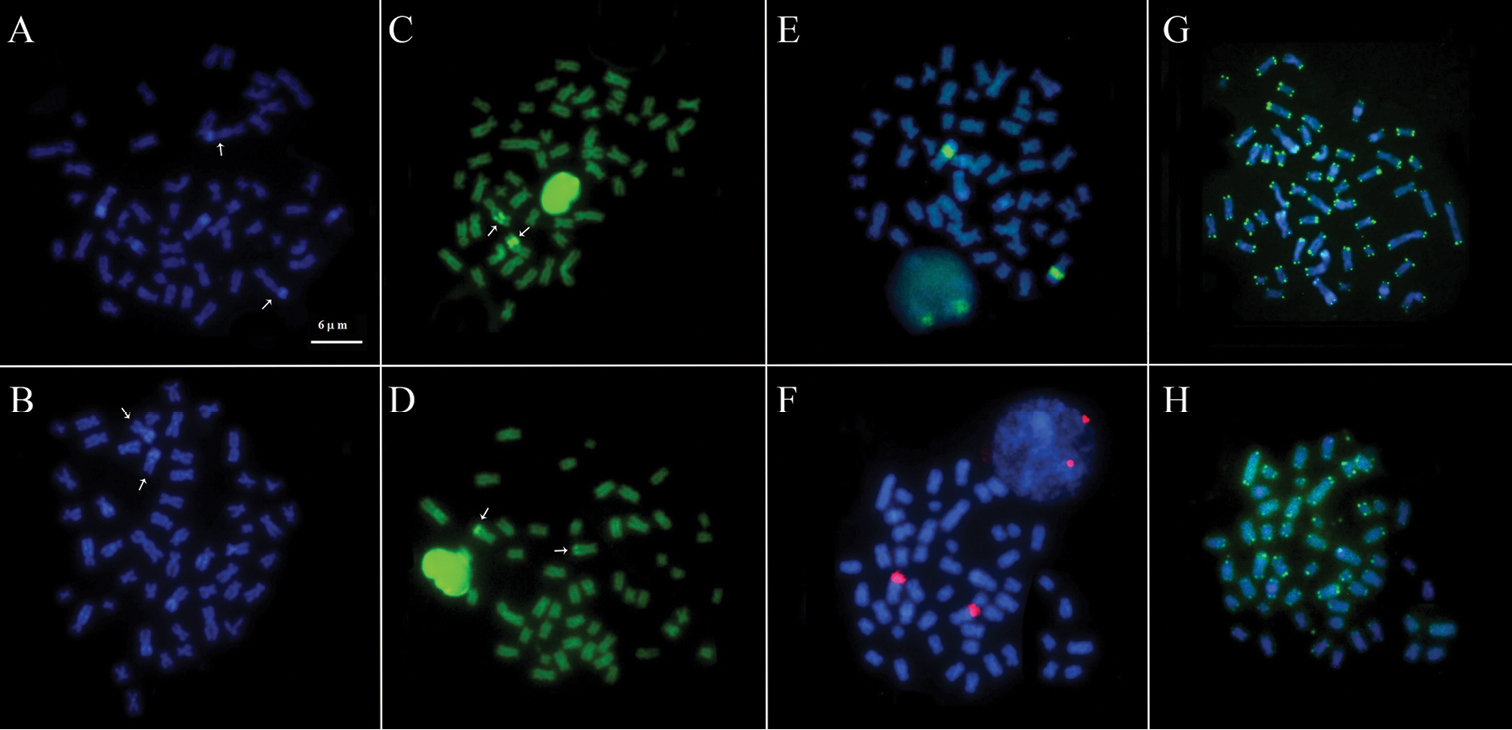
Further karyotypic analyses of Scobinancistrus aureatus (A, C, E and G) and Scobinancistrus pariolispos (B, D, F and H): DAPI staining (A and B), CMA3 staining (C and D), FISH with 18S rDNA probes (E and F) and FISH with telomeric sequence probes (G and H). The scale bar refers to all images.
Scobinancitrus aureatus and Scobinancistrus pariolispos from the Xingu River were found to have the same diploid number (2n=52), but their KFs differed. This is consistent with most other species of tribe Ancistrini, which share 2n=52 and differ in their KFs (
The NOR was found on the same chromosome pair in the two Scobinancistrus species studied here in this situation is shared by most members of the tribe Ancistrini (
The constitutive heterochromatin stained positive with DAPI, indicating that it is AT-rich. Its patterns differed between the two species, with large heterochromatic blocks in non-centromeric regions located predominantly in non-homologous chromosomes, indicating that processes related to the dynamics of repetitive DNA (e.g., transposition) may have been involved in the karyotypic differentiation of these two species.
Using information on karyotype macrostructures and CH and NOR distribution,
In conclusion, the karyotypic differences found in the two Scobinancistrus species studied herein can be used in their taxonomic identification. Moreover, the sympatric occurrence of these species suggests that the identified karyotypic differences may have functioned as a mechanism of post-zygotic reproductive isolation during the speciation process.
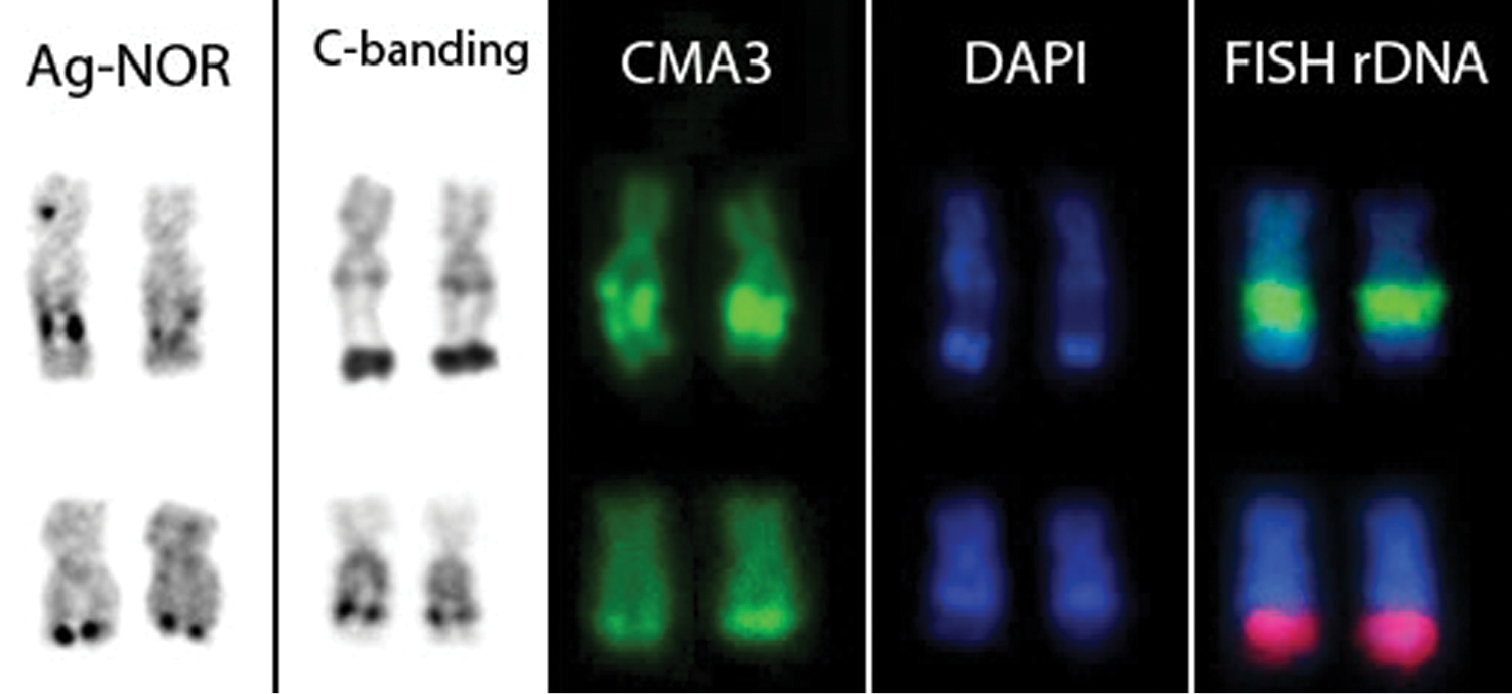
NOR-bearing chromosomes of Scobinancistrus aureatus (upper row) and Scobinancistrus pariolispos (lower row).
We wish to thank Dr. Leandro Melo de Sousa (Federal University of Pará - Campus Altamira) for the taxonomic identification of the samples. Grants to support this research were provided by UFPA, CNPq, CAPES and by the project Vale-FAPESPA. The field collections were authorized by IBAMA (Brazilian Institute of Environment and Renewable Natural Resources).