





(C) 2013 Catarina Nabais. This is an open access article distributed under the terms of the Creative Commons Attribution License 3.0 (CC-BY), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
For reference, use of the paginated PDF or printed version of this article is recommended.
In this study, the description of the karyotypes of the endangered chubs Squalius aradensis (Coelho, Bogutskaya, Rodrigues and Collares-Pereira, 1998) and Squalius torgalensis (Coelho, Bogutskaya, Rodrigues and Collares-Pereira, 1998) is presented by means of conventional (Giemsa-staining, Chromomycin A3 (CMA3)-fluorescence, Silver-impregnation (Ag-NORs)) and molecular (fluorescence in situ hybridization (FISH) with 18S rDNA probe) protocols. These endemic sister-species have an allopatric but adjacent distribution in the most southwestern part of the Iberian Peninsula. Diploid chromosome number was invariably 2n = 50 and karyotypes of both species were grossly similar, composed of metacentric and submetacentric elements with a reduced number of acrocentric pairs. Sequential staining using FISH with an 18S rDNA probe, CMA3 and Ag-NORs treatments revealed consistent positive signals located at the end of the short arms of a submetacentric chromosome pair, likely homologous in both species. While providing useful cytogenetic comparative data against other members of the genus Squalius Bonaparte, 1837, the work aimed to draw attention towards the conservation of two narrow-range and highly confined fish species.
Leuciscinae, Cytotaxonomy, FISH with rDNA, NOR-phenotype, silver staining, chromomycin A3
The genus Squalius Bonaparte, 1837 belongs to the subfamily Leuciscinae, the major element of the Iberian cyprinid fauna. In Portuguese inland waters, four bisexual species with an allopatric distribution (
Most chubs of the genus Squalius were formerly included in the genus Leuciscus Cuvier, 1816 (see
Some cytogenetic data were recorded for populations from Arade, Aljezur and Bordeira drainages (currently assigned to Squalius aradensis) and Mira (currently assigned to Squalius torgalensis) (Collares Pereira et al. 1998), but previously to the acknowledgement that they should be considered distinct taxa from Squalius pyrenaicus (
Adult specimens were captured by electrofishing in two distinct southern populations. Six Squalius aradensis (four males and two females) were collected in the Arade basin (Odelouca River) and two Squalius torgalensis (unknown sex) in the Mira basin (Torgal River). All procedures were developed in accordance to the recommended ethic guidelines (
Chromosome spreads were obtained for one specimen using standard kidney protocol and for the remaining using fin fibroblast cultures (
Chromosomes were stained with a solution of 4% Giemsa (pH=6.8). CMA3 fluorescence staining was performed according to
The physical mapping of major rDNA gene cluster on the chromosomes was accomplished by FISH with rDNA probe. An rDNA clone containing 18S-5.8S-28S genes plus the intergenic spacers and untranscribed sequences from the genome of Drosophila melanogaster Meigen, 1830 (clone pDm 238,
Slides were screened in an Olympus BX 60 epifluorescence microscope equipped with a DP50 Olympus CCD camera. All images were processed using Adobe Photoshop CS4 software. Chromosomes were arranged in a decreasing size order and classified according to their arm ratios (
All Squalius aradensis and Squalius torgalensis karyotypes revealed a diploid number of 2n = 50 chromosomes.
Karyotypes of both Squalius aradensis and Squalius torgalensis are composed of five pairs of metacentric (m), eighteen pairs of submetacentric (sm) and two pairs of subtelo/accrocentric (st/a) chromosomes. As the general karyotypes are the same for both species, only the results for Squalius aradensis using giemsa staining were included in Fig. 1. This genome composition leads to a high fundamental number (NF=96). As regards the eventual occurrence of heteromorphic sex chromosomes, no clear distinction between male and female karyotypes was observed in Squalius aradensis, the species where the specimens’ sex could be accurately assessed.
The NORs’ phenotype was constant throughout all treatments, consistently positively labeling only one NORs-bearing chromosome pair likely homologous in both Squalius aradensis and Squalius torgalensis species. Their rDNA-positive signals were co-localized to CMA3- and Ag-positive signals: in the short arm of a middle-size sm chromosome pair in both species as documented by sequential staining (Figs 2 and 3), indicative of being GC-rich and transcriptional active. No evidences of multi-chromosomal positive NOR’s signals were registered.
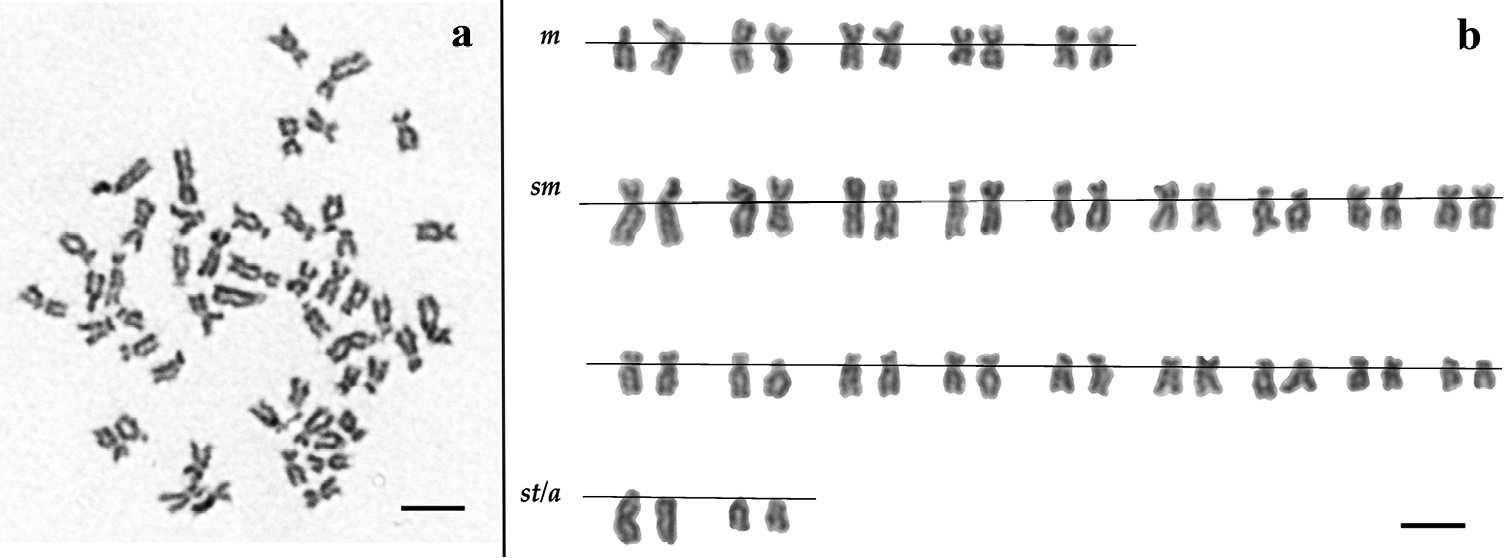
a Giemsa stained metaphase and b corresponding karyotype of a Squalius aradensis male from Odelouca River (Arade drainage). Scale bar = 10µm.
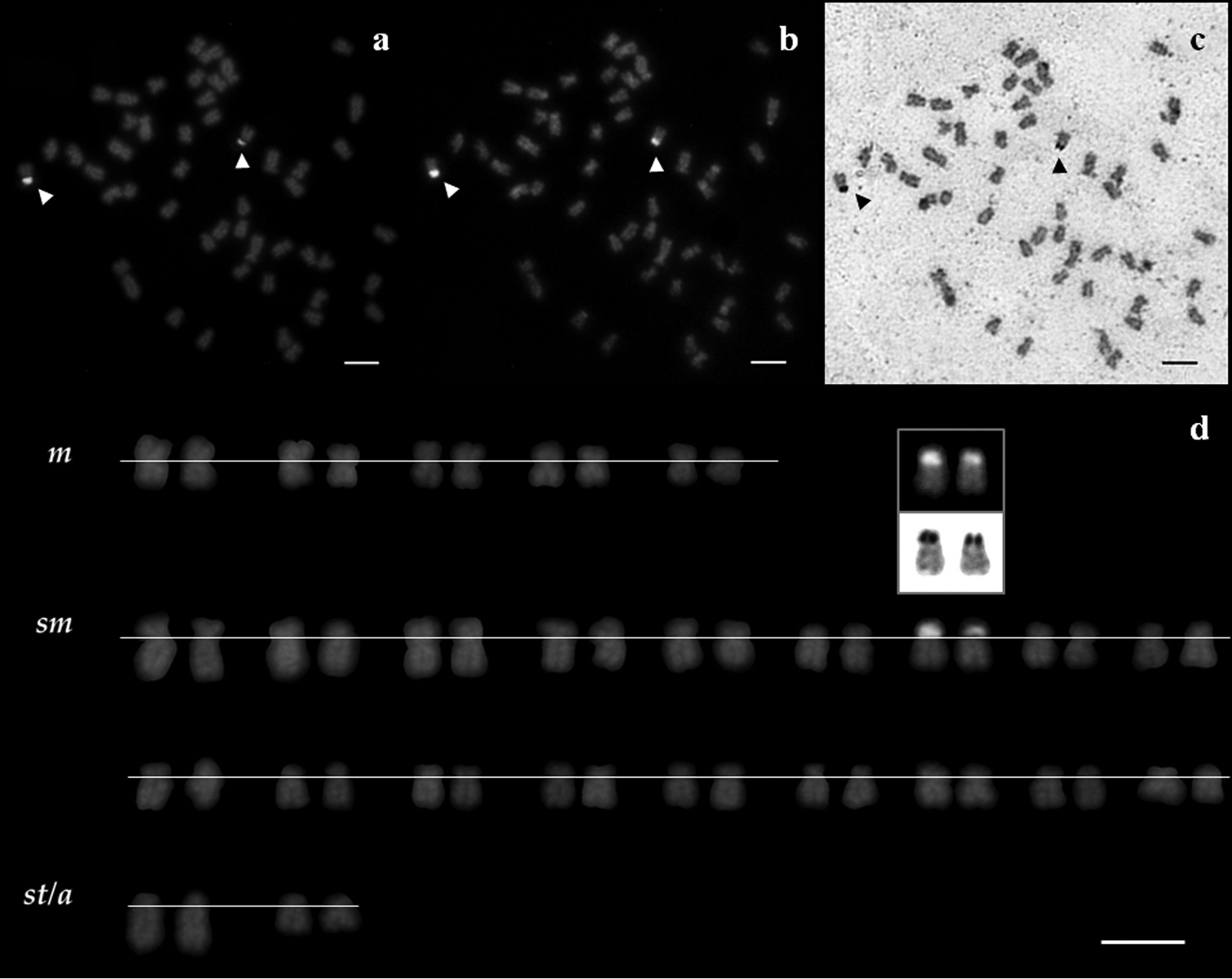
Metaphase spreads of a Squalius aradensis male, stained with sequential treatments a rDNA FISH b CMA3-NOR, and c Ag-NOR treatments d corresponding karyotype of the same individual after rDNA FISH, with CMA3- and Ag-NOR signals inset. NORs are indicated by arrowheads. Scale bar = 10µm.
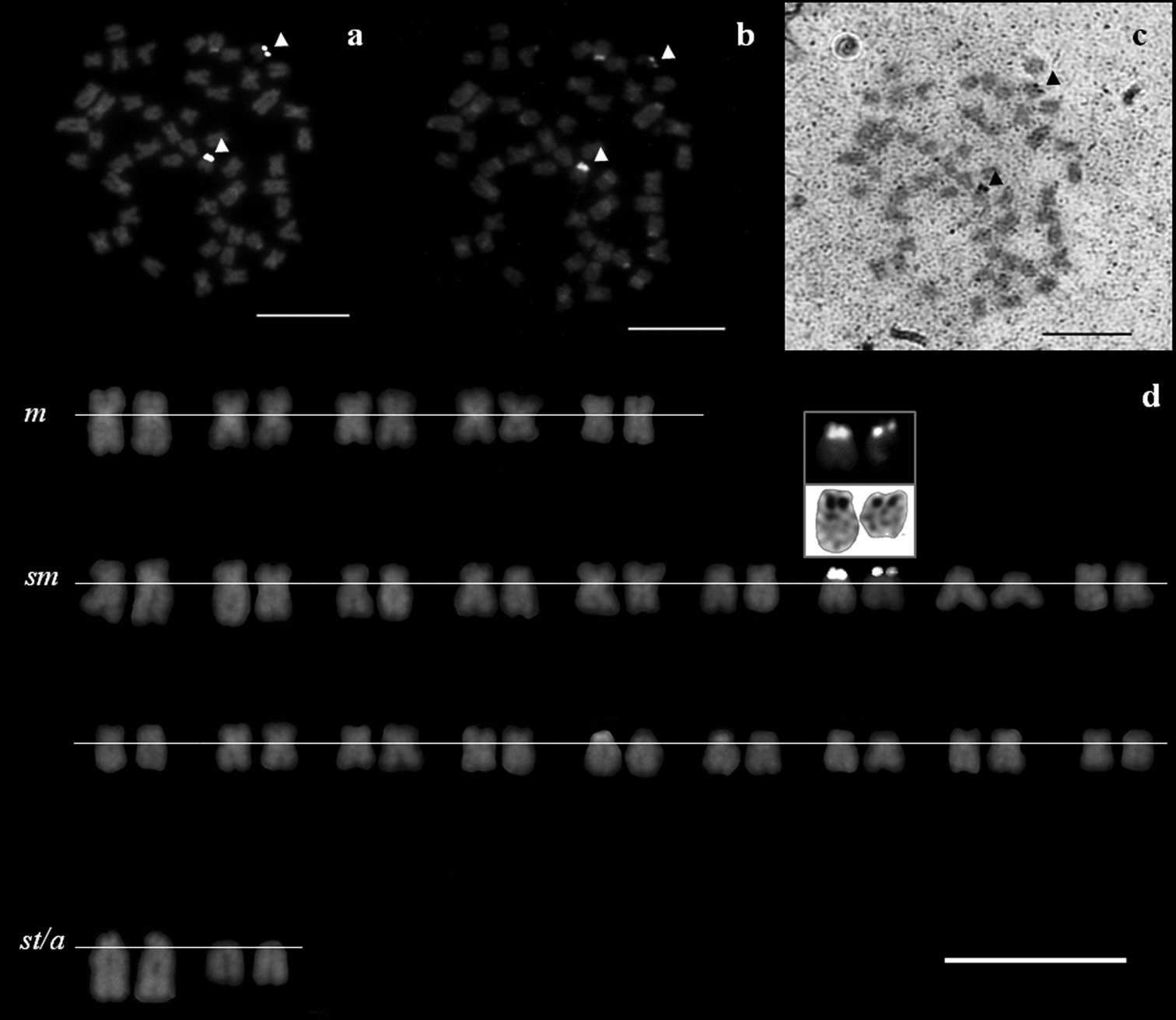
Metaphase spreads of a Squalius torgalensis, stained with sequential treatments a rDNA FISH b CMA3-NOR, and c Ag-NOR treatments d corresponding karyotype of the same individual after rDNA FISH, with CMA3- and Ag-NOR signals inset. NORs are indicated by arrowheads. Scale bar = 10µm.
Squalius aradensis and Squalius torgalensis karyotypes display the general pattern described for most Leuciscinae: a diploid chromosome number of 2n = 50, a chromosome set with mainly bi-armed elements (dominated by submetacentrics with some metacentrics), and only one NOR-bearing chromosome pair. In cyprinids this is assumed to correspond to the ancestral character state and most likely represents also the synapomorphy for the genus Squalius (
The number and location of rDNA gene clusters have been used as chromosome markers in fish cytotaxonomy (e.g.
The karyotypes of the two species proved to be grossly similar at a macrostructural level. They are sister-taxa strongly clustered in many phylogenetic analyses, however consistently reciprocally monophyletic (
Despite the apparent conservative pattern found in the two species here addressed by conventional cytogenetic tools, the karyotype variability present in the Iberian species of the genus Squalius so far analysed (
We thank Carla Pereira for her help during lab work and the remaining CBA Evolutionary Genetics Group in particular Luis Moreira da Costa for assistance in the field, as well as AFN and ICNB for fishing permits. This research was funded by Centro de Biologia Ambiental [PEst-OE/BIA/UI0329/2011]. MR was supported by the FCT Post-Doctoral grant SFRH/BPD/48428/2008, and by projects RPP JU No. CZ.1.07/2.3.00/30.0049 and CENAKVA No. CZ.1.05/2.1.00/01.0024 since November 2012.