Research Article |
|
Corresponding author: Leila Mahiddine-Aoudjit ( mahiddinel1@gmail.com ) Academic editor: Svetlana Galkina
© 2019 Leila Mahiddine-Aoudjit, Ouahida Boucekkine, Kafia Ladjali-Mohammedi.
This is an open access article distributed under the terms of the Creative Commons Attribution License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Citation:
Mahiddine-Aoudjit L, Boucekkine O, Ladjali-Mohammedi K (2019) Banding cytogenetics of the vulnerable species Houbara bustard (Otidiformes) and comparative analysis with the Domestic fowl. Comparative Cytogenetics 13(1): 1-17. https://doi.org/10.3897/CompCytogen.v13i1.30660
|
Abstract
The Houbara bustard Chlamydotis undulata (Jacquin, 1784) is an emblematic and endangered bird of steppes and desert spaces of North Africa. This species belonging to Otidiformes is recognized as vulnerable by the International Union for Nature Conservation.
The critical situation of this species and the revision of its classification on the tree of birds encouraged the authors to start accumulating chromosome data. For that, we propose the GTG- and RBG-banded karyotypes of the Houbara bustard prepared from primary fibroblast cell cultures. The first eight autosomal pairs and sex chromosomes have been described and compared to those of the domestic fowl Gallus domesticus (Linnaeus, 1758). The diploid number has been estimated as 78 chromosomes with 8 macrochromosomes pairs and 30 microchromosomes pairs, attesting of the stability of chromosome number in avian karyotypes.
The description of the karyotype of the Houbara is of crucial importance for the management of the reproduction of this species in captivity. It can be used as a reference in the detection of chromosomal abnormalities, which would be responsible of the early embryonic mortalities.
Keywords
Chlamydotis undulata undulata , endangered species, GTG- and RBG- banded karyotypes, interspecific comparison
Introduction
With approximately 10,699 species, birds represent the class of Tetrapoda with the highest number of species (http://www.worldbirdnames.org). This class presents a certain number of particularities such as the presence of feathers, flight and a small genome (about 1.45 pg that represents 1/3 of the mammalian genome). As well, the avian karyotypes are very particular, with a very consistent diploid number. The range of variation is very wide, between 40 and 138 chromosomes, the average being from 76 to 82 for most species (
Despite the diploid number that seems to be stable in birds, the avian genome has undergone multiple evolutionary events. Chromosome fission has previously been reported as being a factor of evolutionary change (
Besides, phylogenetic analysis of 48 bird species representing all Neoaves orders was conducted and the analysis identified a first divergence of the Neoaves into two independent lines named Passerea and Columbea, without forgetting the emergence of the new order Otidiformes to which the endangered species Houbara bustard is now affiliated (
The Houbara bustard is an emblematic bird of the large steppe areas and desert spaces of North Africa and the Middle East (
No description of the karyotype of the Houbara bustard has been reported to date. The only known cytogenetic data are a metaphase of this species, without a precise description of the chromosomes, which was reported in the study that allowed the development of chromosome paints and BACs for the characterization of inter- and intrachromosomal rearrangements of avian microchromosomes (
Here we describe the macrochromosomes of the Houbara bustard in morphologic GTG bands and dynamic RBG bands. Morphometric measurements were used to facilitate the classification of smaller macrochromosomes. The obtained banding pattern in Houbara bustard chromosomes was compared with that of the chicken chromosomes, in order to determine the presence of chromosomal rearrangements that would have occurred during speciation.
Material and methods
Embryos
Fifteen Houbara bustard embryos aged between 8 and 19 days were collected from Emirati Bird Breeding Centre for Conservation EBBCC (32°55'40.54"N, 0°32'33.71"E) in the region of Abiod Sidi Cheikh (Wilaya d’El-Bayadh, south of Algeria) during the breeding season. The embryos were obtained in accordance with the authorization from the General Direction of Forests of Algeria (N°30BOG/N°80DPFF/DGF-18).
Cell culture and chromosomes preparations
Fibroblasts were isolated from Houbara bustard embryos by trypsinisation (trypsin solution 0.05%, Sigma) and incubated at 41 °C with RPMI 1640 culture medium (20 mM HEPES, Gibco) supplemented with 10% of fetal calf serum (FCS) (Gibco), 1% L-Glutamine (Sigma), 1% penicillin, streptomycin and fungizone (Sigma). Cultures of fibroblasts were synchronised as described by
As a sufficient number of refractive mitotic cells was observed (after 6–8 h), they were treated with colchicine (final concentration: 0.05 μg/ml, Sigma) for 5 min at 37 °C. Cells were harvested by the addition of 0.05% trypsin-EDTA (Gibco). Hypotonic treatment was performed. In fact, cells were suspended for 13 min at 37 °C in hypotonic solution 1:5 (FCS- distilled water). Fixation and spreading were performed using standard methods (
Staining procedures
The revelation of the structural GTG bands is based on enzymatic digestion with proteolysis (
The RBG-FPG staining (R-bands obtained with BrdU by Fluorochrome-photolysis and Giemsa staining) was performed as previously described (
To make a comparison with chromosomes of the chicken, GTG banding was also performed on previously frozen chicken chromosome preparations.
Chromosome classification and measurement
Slides were analysed using Axio Scope A1 (Zeiss) and thirty metaphases with decondensed chromosomes were selected and photographed with CoolCube1 (Metasystems). Houbara bustard chromosomes were classified according to the International System of Standardised Avian Karyotypes (ISSAK) (
The first eight pairs of chromosomes of the Houbara bustard and the domestic fowl and their sex chromosomes were measured using KARYOTYPE 2.0 software (
Results
The diploid number of the Houbara bustard has been estimated as 78 chromosomes by examination of full metaphases (Fig.

Metaphase of the Houbara bustard showing macrochromosomes and microchromosomes with Giemsa staining. Scale bar: 5 μm.
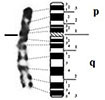
In this study, we propose the karyotype of the Houbara bustard with morphological GTG-banded chromosomes (Fig.

GTG (A) and RBG (B) karyotypes of the first 12 and sex chromosomes of Houbara bustard Chlamydotis undulata undulata. Scale bar: 5 μm.

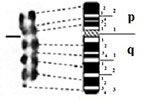
GTG-banded (A) and RBG-banded (B) macrochromosomes, sex chromosomes ZW and their corresponding ideograms of Houbara bustard Chlamydotis undulata undulata.
Description of GTG and RBG bands on macrochromosomes and sex chromosomes ZW of the Houbara bustard.
| Chromosomes | GTG bands / ideograms | Description | RBG bands / ideograms | Description |
|---|---|---|---|---|
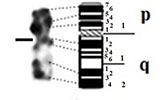
| 1 |
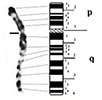
|
– Short arm (p) |
|
– Short arm (p) |
| submetacentric | 2 regions | 2 regions | ||
| 9 bands | 7 bands | |||
| – Long arm (q) | – Long arm (q) | |||
| 4 regions | 4 regions | |||
| 16 bands | 15 bands | |||
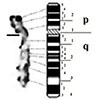
| 2 |
|
– Short arm (p) |
|
– Short arm (p) |
| submetacentric | 3 regions | 2 regions | ||
| 8 bands | 6 bands | |||
| – Long arm (q) | – Long arm (q) | |||
| 3 regions | 3 regions | |||
| 13 bands | 10 bands | |||
| 3 |
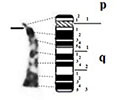
|
– Short arm (p) |
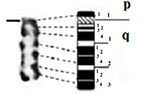
|
– Short arm (p) |
| acrocentric | 1 region | 1 region | ||
| 2 bands | 1 band | |||
| – Long arm (q) | – Long arm (q) | |||
| 3 regions | 3 regions | |||
| 14 bands | 12 bands | |||
| 4 |

|
– Short arm (p) |
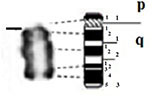
|
– Short arm (p) |
| acrocentric | 1 region | 1 region | ||
| 1 band | 1 band | |||
| – Long arm (q) | – Long arm (q) | |||
| 3 regions | 3 regions | |||
| 11 bands | 9 bands | |||
| 5 |
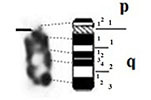
|
– Short arm (p) |
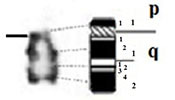
|
– Short arm (p) |
| acrocentric | 1 regions | 1 regions | ||
| 2 bands | 1 band | |||
| – Long arm (q) | – Long arm (q) | |||
| 3 regions | 2 regions | |||
| 7 bands | 6 bands | |||
| 6 |
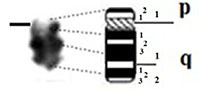
|
– Short arm (p) |
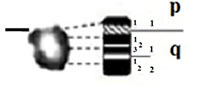
|
– Short arm (p) |
| acrocentric | 1 region | 1 region | ||
| 2 bands | 1 band | |||
| – Long arm (q) | – Long arm (q) | |||
| 2 regions | 2 regions | |||
| 6 bands | 5 bands | |||
| 7 |
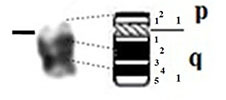
|
– Short arm (p) |
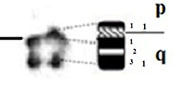
|
– Short arm (p) |
| acrocentric | 1 region | 1 region | ||
| 2 bands | 1 band | |||
| – Long arm (q) | – Long arm (q) | |||
| 1 region | 1 region | |||
| 5 bands | 3 bands | |||
| 8 |
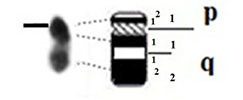
|
– Short arm (p) |
|
– Short arm (p) |
| acrocentric | 1 region | 1 region | ||
| 2 bands | 1 band | |||
| – Long arm (q) | – Long arm (q) | |||
| 2 regions | 1 region | |||
| 3 bands | 3 bands | |||
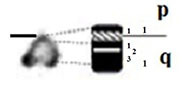
| Z |
|
– Short arm (p) |
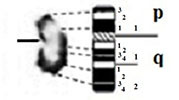
|
– Short arm (p) |
| submetacentric | 1 region | 1 region | ||
| 7 bands | 3 bands | |||
| – Long arm (q) | – Long arm (q) | |||
| 2 regions | 2 regions | |||
| 10 bands | 8 bands | |||
| W |
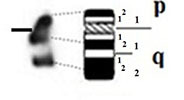
|
- Short arm (p) |
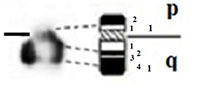
|
- Short arm (p) |
| submetacentric | 1 regions | 1 regions | ||
| 2 bands | 2 bands | |||
| - Long arm (q) | - Long arm (q) | |||
| 2 regions | 2 regions | |||
| 4 bands | 4 bands |
The haploid karyotype of the first 10 autosomes and the sex chromosomes Z and W of the Houbara bustard corresponds to 130 GTG bands and 104 RBG bands. The number of bands obtained for this species is lower than that reported in the chicken for the same chromosome number (209 bands G and 182 bands R) (
In order to compare the chromosomes of the Houbara bustard and those of the domestic fowl, the first eight macrochromosomes and the sex chromosomes ZW of these two species have been measured (Table
Measurements of eight macrochromosomes and sex chromosomes ZW of Houbara bustard Chlamydotis undulata undulata and Domestic fowl Gallus domesticus. Chr: chromosome, q: long arm, p: short arm, p+q: total length, r: arm ratio q/p, CI %: centromeric index=p/p+q × 100. Lengths are given in micrometer (µm) ± standard deviation.
| Chr | Houbara bustard | Domestic fowl | ||||||||
| q | p | p+q | r | CI % | q | p | p+q | r | CI % | |
| 1 | 2.93±0.57 | 1.19±0.29 | 4.12±0.81 | 2.46 | 29 % | 6.69±1.26 | 3.97±0.70 | 10.66±1.85 | 1.69 | 37% |
| 2 | 2.08±0.41 | 0.95±0.24 | 3.02±0.63 | 2.19 | 31 % | 5.35±0.71 | 2.76±0.54 | 8.11±1.24 | 1.94 | 34% |
| 3 | 2.15±0.47 | 0.12±0.05 | 2.26±0.49 | 18.50 | 5 % | 5.42±0.66 | 0.36±0.05 | 5.77±0.62 | 15.18 | 6% |
| 4 | 1.33±0.25 | 0.12±0.05 | 1.45±0.27 | 10.98 | 8 % | 3.72±0.59 | 0.96±0.15 | 4.69±0.72 | 3.86 | 20% |
| 5 | 1.21±0.20 | 0.09±0.04 | 1.29±0.22 | 13.37 | 7 % | 2.71±0.43 | 0.29±0.12 | 3.00±0.51 | 9.39 | 9% |
| 6 | 0.87±0.16 | 0.06±0.06 | 0.92±0.18 | 15.86 | 6 % | 1.53±0.16 | 0.07±0.10 | 1.60±0.24 | 21.83 | 4% |
| 7 | 0.79±0.11 | 0.02±0.03 | 0.77±0.12 | 41.89 | 2 % | 1.28±0.23 | 0.40±0.05 | 1.68±0.28 | 3.18 | 24% |
| 8 | 0.67±0.13 | 0.01±0.02 | 0.67±0.12 | 92.52 | 1 % | 0.89±0.14 | 0.61±0.05 | 1.49±0.15 | 1.46 | 41% |
| Z | 0.99±0.20 | 0.46±0.13 | 1.44±0.31 | 2.17 | 32 % | 2.30±0.34 | 2.04±0.29 | 4.34±0.63 | 1.12 | 47% |
| W | 0.59±0.15 | 0.20±0.07 | 0.78±0.17 | 3.01 | 25 % | 0.95±0.35 | 0.60±0.28 | 1.55±0.60 | 1.59 | 39% |
Discussion
The diploid number has been estimated in the Houbara bustard as 78 chromosomes as in many birds. Indeed, the diploid number is highly conserved with about 63% of birds with a chromosome number that varies between 74 and 86 (
The size of the first eight pairs of chromosomes of the Houbara bustard varies between 4 μm (chromosome 1) and 0.67 μm (chromosome 8). This average size of Houbara bustard macrochromosomes is lower to the estimated size (3 to 6 μm) for avian macrochromosomes (
The comparison of the first eight pairs and sex chromosomes of the Houbara bustard with those of the chicken revealed the presence of similarities as well as differences between these two species. Indeed the karyotype of the chicken conserves the ancestral karyotype of many avian orders (
The first three chromosomes of the Houbara bustard are morphologically similar to those of the chicken. Chromosome 1 and 2 are submetacentric and chromosome 3 is acrocentric. These results are in agreement with those of
The comparison of the chromosome 1 of Houbara bustard with that of the chicken revealed a difference in the size of the p- arm of chromosome 1 (1p) which is shorter in the bustard (Fig.

Representation of chromosomal rearrangements that could have occurred during the formation of the chromosome 1 (A), the chromosome 2 (B), the chromosome 4 (C) and the chromosome Z (D) of the Houbara bustard.
As for chromosomes 1 of Japanese quail and chicken, the high-resolution mapping using fluorescent in situ hybridisation with molecular markers on stretched chromosomes in the lampbrush showed that the difference in the morphology of this chromosome should be explained by de novo centromere formation and the hypothesis of centrometric inversion should be excluded (
Likewise, the long arm (q) of chromosome 2 of the Houbara bustard is shorter than that of the chicken and that would be the consequence of a terminal fission (Fig.
The chromosome 4 of the Houbara bustard is acrocentric while that of the chicken is telocentric. Their arm ratios are equal to 10.98 and 3.86 respectively (Table
The chromosome 5 of the Houbara bustard is acrocentric like that of the chicken but it seems to have lost the terminal part of the long arm. Indeed, chromosome 5 of the Houbara bustard measures 1.29 ± 0.22 µm and that of the chicken 3 ± 0.51 µm. This could be explained by a fission event that would have occurred during evolution. In fact, chromosome 5 of the chicken appears to be distributed on the short arm (p) of chromosome 4 of the coot (FAT4) and on microchromosome 12 (FAT12) (
In contrast to chromosome 6 of the bustard which appears to be morphologically similar to that of the chicken, the chromosomes 7 and 8 of these two species are different. Houbara bustard chromosomes 7 and 8 are acrocentric whereas they are, respectively, telocentric and submetacentric in chicken (
Finally, the sex chromosome Z of the Houbara bustard differs from that of the chicken. It is submetacentric in the first species and metacentric in the second. The arm ratio (q/p) is 2.17 for the chromosome Z of the bustard while it is equal to 1.12 for that of the chicken (Table
Chromosome Z that is metacentric in chicken appears to be submetacentric in many other species of Galliformes (
Despite the conservation of this chromosome in its totality during evolution, it appears to be subject to intrachromosomal rearrangements (
The W chromosome is submetacentric in the Houbara bustard. It appears to be morphologically similar to that of the chicken. Depending on its size, it can be classified between chromosome 6 and 7. The W chromosome in birds and reptiles seems to have degenerated during evolution. It is physically small, with a high proportion of heterochromatin (
In conclusion, this analysis of the chromosomes of the endangered Houbara bustard provided a precise description of a part of its karyotype in GTG and RBG bands. Chromosomal informations have been obtained for the newly established Otidiformes order. The identification of microchromosomes by fluorescence in situ hybridisation of specific BAC clones of chicken chromosomes is conceivable to complete the description of the karyotype of this species. The various rearrangements suggested must be confirmed by molecular studies of BAC clones localisation and chromosome painting for a better knowledge of avian karyotypes evolution.
Acknowledgements
The present work has received a financial assistance from:
– Ministère de l’Aménagement du Territoire et de l’Environnement in the framework of the project n° 223 : «Stratégie d’études pour la conservation et la valorisation de la biodiversité du Sud algérien : l’outarde houbara Chlamydotis undulata».
– Ministère de l’Enseignement Supérieur et de la Recherche Scientifique in the framework of the national project n° 209 «Analyse génomique de trois espèces d’oiseaux menacées de disparition».
Special thanks to: Berahal M. for her technical support, Pr. Mohammedi A. (University M’Hamed Bougara of Boumerdes, Algeria), Pr. Belhamra M. (University of Biskra, Algeria) and Mr. Chebouti Y. (INRF) for the interest they exhibit to this work. We would like to thank our collaborators from the forest conservation of El-Bayadh and those of EBBC Center for providing biological material, especially Mr. Scotland K., Mr. Bouabdellah K., Mr. Madani M. and Mr. Amari T. Without forgetting, AJZ Engineering Algeria (Zeiss company), in particular Mr. Laguel F. for his precious help.
References
- Altinordu F, Peruzzi L, Yu Y, He X (2016) A tool for the analysis of chromosomes: KaryoType. Taxon 65(3): 586–592. https://doi.org/10.12705/653.9
- Arif IA, Khan HA, Williams JB, Shobrak M, Arif WI (2012) DNA Barcodes of Asian Houbara Bustard (Chlamydotis undulata macqueenii). International Journal of Molecular Sciences 13: 2425–2438. http://doi.org/10.3390/ijms13022425
- Auer H, Mayr B, Lambrou M, Schleger W (1987) An extended chicken karyotype, including the NOR chromosome. Cytogenetic and Genome Research 45(3–4): 218–221. https://doi.org/10.1159/000132457
- Azafzaf H, Sande E, Evans SW, Smart M, Collar NJ (2005) International Action plan for North African Houbara Bustard Chlamydotis undulata undulata. Birdlife International, Nairobi, Kenya and Royal Society for the Protection of Birds, Sandy, 31 pp.
- Belterman RHR, De Boer LEM (1984) A karyological study of 55 species of birds including karyotypes of 39 species new to cytology. Genetica 65(1): 39–82. https://doi.org/10.1007/BF00056765
- Chbel F, Broderick D, Idaghdour Y, Korrida A, McCormick P (2002) Characterization of 22 microsatellites loci from the endangered Houbara bustard (Chlamydotis undulata undulata). Molecular Ecology Notes 2(4): 484–487. https://doi.org/10.1046/j.1471-8286.2002.00287.x
- Christidis L (1990) Animal cytogenetics Chordata 3B: Aves. Stuttgart, 116 pp.
- Cracraft J (1981) Toward a phylogenetic classification of the recent birds of the world (Class Aves). Auk 98: 681–714.
- De Boer LEM (1976) The somatic chromosome complements of 16 species of Falconiformes (Aves) and the karyological relationships of the order. Genetica 46: 77–113. https://doi.org/10.1007/BF00122519
- De Boer LEM, Sinoo RP (1984) A karyological study of Accipitridae (Aves: Falconiformes), with karyotypic descriptions of 16 species new to cytology. Genetica 65(1): 89–107. https://doi.org/10.1007/BF00056767
- Del Hoyo J, Collar NJ, Christie DA, Elliott A, Fishpool LDC (2014) HBW and BirdLife International, Illustrated Checklist of the Birds of the World. Volume 1: Non-passerines (Eds) Lynx, BirdLife International, Barcelona–Cambridge, 904 pp.
- Dutrillaux B, Couturier J (1981) La pratique de l’analyse chromosomique (Eds) Masson, Paris, 84 pp.
- Ellegren H (2011) Sex-chromosome evolution: recent progress and the influence of male and female heterogamety. Nature Reviews Genetics 12: 157–166. https://doi.org/10.1038/nrg2948
- Fain MG, Houde P (2004) Parallel radiations in the primary clades of birds. Evolution 58(11): 2558–2573. https://doi.org/10.1111/j.0014-3820.2004.tb00884.x
- Furo IDO, Monte A, Santos M, Tagliarini MM, O´Brien P, Ferguson-Smith MA, De Oliveira EHC (2015) Cytotaxonomy of Eurypyga helias (Gruiformes, Eurypygidae): First karyotypic description and phylogenetic proximity with Rynochetidae. PlosOne 12: e0147458. https://doi.org/10.1371/journal.pone.0143982
- Gray JE (1832) Otis macqueenii – Hardwicke’s illustration. Indian Zoology 2, London, 220 pp.
- Griffin DK, Robertson LB, Tempest HG, Skinner BM (2007) The evolution of the avian genome as revealed by comparative molecular cytogenetics. Cytogenetic and Genome Research 117(1–4): 64–77. https://doi.org/10.1159/000103166
- Guttenbach M, Nanda I, Feichtinger W, Masabanda JS, Griffin DK, Schmid M (2003) Comparative chromosome painting of chicken autosomal paints 1–9 in nine different bird species. Cytogenetic and Genome Research 103(1–2): 173–184. https://doi.org/10.1159/000076309
- Heim de Balsac H, Mayaud N (1962) Les Oiseaux du Nord-Ouest de l’Afrique. Paul le chevalier, Paris, 448 pp.
- Houde P, Cooper A, Leslie E, Strand AE, Montano GA (1997) Phylogeny and evolution of 12S rDNA in Gruiformes (Aves). In: Mindell DP (Ed.) Avian molecular evolution and systematics, New York, 117–154. https://doi.org/10.1016/B978-012498315-1/50009-1
- Jacquin JF (1784) Psophia undulata. Beyträge zur Geschichte der Vögel 24: 1–9.
- Jarvis ED, Mirarab S, Aberer AJ, Li B, Houde P, Li C, Ho SY, Faircloth BC, Nabholz B, Howard JT, Suh A, Weber CC, Fonseca DD, Li J, Zhang F, Li H, Zhou L, Narula N, Liu L, Ganapathy G, Boussau B, Bayzid MS, Zavidovych V, Subramanian S, Gabaldón T, Capella-Gutiérrez S, Huerta-Cepas J, Rekepalli B, Munch K, Schierup M, Lindow B, Warren WC, Ray D, Green RE, Bruford MW, Zhan X, Dixon A, Li S, Li N, Huang Y, Derryberry EP, Bertelsen MF, Sheldon FH, Brumfield RT, Mello CV, Lovell PV, Wirthlin M, Schneider MPC, Prosdocimi F, Samaniego JA, Velazquez AMV, Alfaro-Núñez A, Campos PF, Petersen B, Sicheritz-Ponten T, Pas A, Bailey T, Scofield P, Bunce M, Lambert DM, Zhou Q, Perelman P, Driskell AC, Shapiro B, Xiong Z, Zeng Y, Liu S, Li Z, Liu B, Wu K, Xiao J, Yinqi X, Zheng Q, Zhang Y, Yang H, Wang J, Smeds L, Rheindt FE, Braun M, Fjeldsa J, Orlando L, Barker FK, Jønsson KA, Johnson W, Koepfli KP, O’Brien S, Haussler D, Ryder OA, Rahbek C, Willerslev E, Graves GR, Glenn TC, McCormack J, Burt D, Ellegren H, Alström P, Edwards SV, Stamatakis A, Mindell DP, Cracraft J, Braun EL, Warnow T, Jun W, Gilbert MTP, Zhang G (2014) Whole-genome analyses resolve early branches in the tree of life of modern birds. Science 346(6215): 1320–1331. https://doi.org/10.1126/science.1253451
- Kretschmer R, De Lima VLC, De Souza MS, Costa AL, O’Brien PCM, Ferguson-Smith MA, De Oliveira EHC, Gunski RJ, Garnero ADV (2018a) Multidirectional chromosome painting in Synallaxis frontalis (Passeriformes, Furnariidae) reveals high chromosomal reorganization, involving fissions and inversions. Comparative Cytogenetics 12(1): 97–110. https://doi.org/10.3897/CompCytogen.v12i1.22344
- Ladjali K, Tixier-Boichard M, Gibiu EP (1995) High resolution chromosome preparations for G- and R- Banding in Gallus domesticus. Journal of Heredity 86: 136–139. https://doi.org/10.1093/oxfordjournals.jhered.a111543
- Ladjali-Mohammedi K, Bitgood JJ, Tixier-Boichard M, Ponce de Leon FA (1999) International System for Standardized Avian Karyotypes (ISSAK): standardized banded karyotypes of the domestic fowl (Gallus domesticus). Cytogenetic and Genome Research 86: 271–276. https://doi.org/10.1159/000015318
- Le Cuziat J, Lacroix F, Roche P, Vidal E, Médail F, Orhant N, Béranger PM (2005) Landscape and human influences on the distribution of the endangered North African houbara bustard (Chlamydotis undulata undulata) in Eastern Morocco. Animal Conservation 8(2): 143–152. https://doi.org/10.1017/S1367943005001903
- Lithgow PE, O’Connor R, Smith D, Fonseka G, Al Mutery A, Rathje C, Frodsham R, O’Brien P, Kasai F, Ferguson-Smith MA, Skinner BM, Griffin DK (2014) Novel tools for characterising inter and intra chromosomal rearrangements in avian microchromosomes. Chromosome Research 22(1): 85–97. https://doi.org/10.1007/s10577-014-9412-1
- Livezey BC (1998) A phylogenetic analysis of the Gruiformes (Aves) based on morphological characters with an emphasis on the rails (Rallidae). Philosophical Transactions of the Royal Society of London B 353: 2077–2151. https://doi.org/10.1098/rstb.1998.0353
- McQueen HA, Fantes J, Cross SH, Clark VH, Archibald AL, Bird AP (1996) CpG islands of chicken are concentrated on microchromosomes. Nature Genetics 12: 321–324. https://doi.org/10.1038/ng039_6-321
- Nanda I, Schlegelmilch K, Haaf T, Schartl M, Schmid M (2008) Synteny conservation of the Z chromosome in 14 avian species (11 families) supports a role for Z dosage in avian sex determination. Cytogenetic and Genome Research 122(2): 150–156. https://doi.org/10.1159/000163092
- Nanda I, Benisch P, Fetting D, Haaf T, Schmid M (2011) Synteny conservation of chicken macrochromosomes 1–10 in different avian lineages revealed by cross-species chromosome painting. Cytogenetic and Genome Research 132(3): 165–181. https://doi.org/10.1159/000322358
- Olson SL (1985) The fossil record of birds. In: Farner DS, King JR, Parkes KC (Eds) Avian Biology, New York, 79–238. https://doi.org/10.1016/B978-0-12-249408-6.50011-X
- Ouchia-Benissad S, Ladjali-Mohammedi K (2018) Banding cytogenetics of the Barbary partridge Alectoris barbara and the Chukar partridge Alectoris chukar (Phasianidae): a large conservation with Domestic fowl Gallus domesticus revealed by high resolution chromosomes. Comparative Cytogenetics 12(2): 171–199. https://doi.org/10.3897/CompCytogen.v12i2.23743
- Perry J, Slater HR, Choo KHA (2004) centric fission simple and complex mechanisms. Chromosome Research 12: 627–640. https://doi.org/10.1023/B:CHRO.0000036594.38997.59
- Pitra C, D’Aloia MA, Lieckfeldt D, Combreau O (2004) Genetic variation across the current range of the Asian Houbara Bustard (Chlamydotis undulata macqueenii). Conservation Genetics 2: 205–215. https://doi.org/10.1023/B:COGE.0000030004.51398.28
- Prum OR, Berv JS, Dornburg A, Field DJ, Towsend JP, Lemmon EM, Lemmon AR (2015) A comprehensive phylogeny of birds (Aves) using targeted next generation DNA sequencing. Nature 526: 569–572. https://doi.org/10.1038/nature15697
- Rodionov V (1996) Micro versus macro: a review of structure and function of avian micro- and macrochromosomes. Russian Journal of Genetics 32: 517–527.
- Rodionov AV (1997) Evolution of avian chromosomes and linkage groups. Russian Journal of Genetics 33: 605–617.
- Romagnano A, Richer CL (1984) R- banding of horse chromosomes: The fluorescence-photolysis-Giemsa technique after bromodeoxyuridine incorporation. Journal of Heredity 75(4): 269–272. https://doi.org/10.1093/oxfordjournals.jhered.a109930
- Roselaar CS (1980) Family Otididae – bustards. In: Cramp S, Simmons KEL (Eds) Handbook of the birds of Europe, the Middle East and North Africa.Vol. 2: Hawks to Bustards. Oxford, 636–668.
- Rothschild W, Hartert E (1894) On a new Bustard from the Palearctic region. Novitates Zoologicae 1: 689.
- Schartl M, Schmid M, Nanda I (2016) Dynamics of vertebrate sex chromosome evolution: from equal size to giants and dwarfs. Chromosoma 125(3): 553–571. https://doi.org/10.1007/s00412-015-0569-y
- Schmid M, Enderle E, Schindler D, Schempp W (1989) Chromosome banding and DNA replication patterns in bird karyotypes. Cytogenetics and Cell Genetics 52(3–4): 139–146. https://doi.org/10.1159/000132864
- Seabright M (1971) A rapid banding technique for human chromosomes. Lancet 2(7731): 971–972. https://doi.org/10.1016/S0140-6736(71)90287-X
- Shibusawa M, Nishibori M, Nishida-Umehara C, Tsudzuki M, Masabanda J, Griffin DK, Matsuda Y (2004) Karyotypic evolution in the Galliformes: an examination of the process of karyotypic evolution by comparison of the molecular cytogenetic findings with the molecular phylogeny. Cytogenetic and Genome Research 106(1): 111–119. https://doi.org/10.1159/000078570.
- Sibley CG, Ahlquist JE (1990) Phylogeny and Classification of Birds: A Study in Molecular Evolution. New Haven, 1080 pp.
- Sibley CG, Ahlquist JE, De Benedictis P (1993) The phylogenetic relationships of the rails, based on DNA comparisons. Journal of the Yamashina Institute for Ornithology 25(1): 1–11. https://doi.org/10.3312/jyio1952.25.1
- Skinner BM, Griffin DK (2012) Intrachromosomal rearrangements in avian genome evolution: evidence for regions prone to breakpoints. Heredity 108(1): 37–41. https://doi.org/10.1038/hdy.2011.99
- Smith J, Bruley CK, Paton IR, Dunn I, Jones CT, Windsor D, Morrice DR, Law AS, Masabanda J, Sazanov A, Waddington D, Fries R, Burt DW (2000) Differences in gene density on chicken macrochromosomes and microchromosomes. Animal Genetics 31(2): 96–103. https://doi.org/10.1046/j.1365-2052.2000.00565.x
- Smith J, Burt DW (1998) Parameters of the chicken genome (Gallus gallus). Animal Genetics 29(4): 290–294. https://doi.org/10.1046/j.1365-2052.1998.00334.x
- Takagi N, Sasaki M (1974) A phylogenetic study of bird karyotypes. Chromosoma 46(1): 91–120. https://doi.org/10.1007/BF00332341
- Tegelström H, Ryttman H (1981) Chromosomes in birds (Aves): evolutionary implications of macro and microchromosomes numbers and lengths. Hereditas 94(2): 225–233. https://doi.org/10.1111/j.1601-5223.1981.tb01757.x
- Tegelström H, Ebenhard T, Ryttman H (1983) Rate of karyotype evolution and speciation in birds. Hereditas 98(2): 235–239. https://doi.org/10.1111/j.1601-5223.1983.tb00600.x
- Tobias JA, Seddon N, Spottiswoode CN, Pilgrim JD, Fishpool LDC, Collar NJ (2010) Quantitative criteria for species delimitation. Ibis 152(4): 724–746. https://doi.org/10.1111/j.1474-919X.2010.01051.x
- Viegas-Péquignot E, Dutrillaux B, Magdelenat H, Coppey-Moisan M (1989) Mapping of a single copy DNA sequences on human chromosomes by in situ hybridization with biotinylated probes: enhancement of detection sensitivity by intensified fluorescence digital imaging microscopy. Proceedings of the National Academy of Sciences USA 86(2): 582–586. https://doi.org/10.1073/pnas.86.2.582
- Wetmore A (1960) A classification for the birds of the world. Smithsonian Miscellaneous Collections 139(11): 1–37.
- Zakharov AF, Egolina NA (1968) Asynchrony of DNA replication and mitotic spiralization along heterochromatic portions of Chinese hamster chromosomes. Chromosoma 23(4): 365–385. https://doi.org/10.1007/BF00625285
- Zlotina A, Galkina S, Krasikova A, Crooijmans RP, Groenen MAM, Gaginskaya E, Deryusheva S (2012) Centromere positions in chicken and Japanese quail chromosomes: de novo centromere formation versus pericentric inversions. Chromosome Research 20(8): 1017–1032. https://doi.org/10.1007/s10577-012-9319-7